

Rosmarinic Acid Present in Lepechinia floribunda and Lepechinia meyenii as a Potent Inhibitor of the Adenylyl Cyclase gNC1 from Giardia lamblia
 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Plants from Central Argentina as Inhibitors of the Adenylyl Cyclase gNC1 from Giardia lamblia
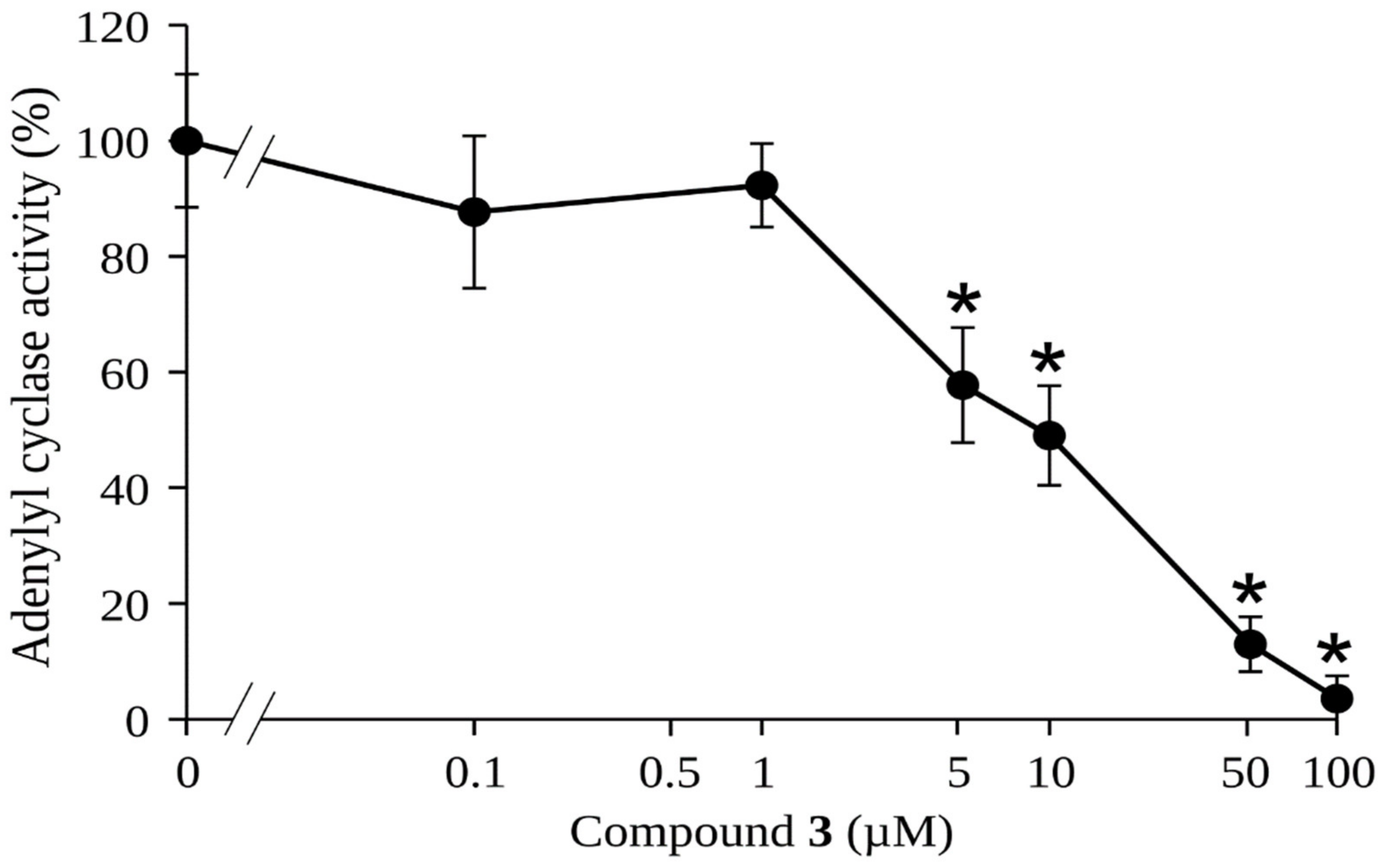
2.2. Effect of Rosmarinic Acid (3) on the Activity of gNC1
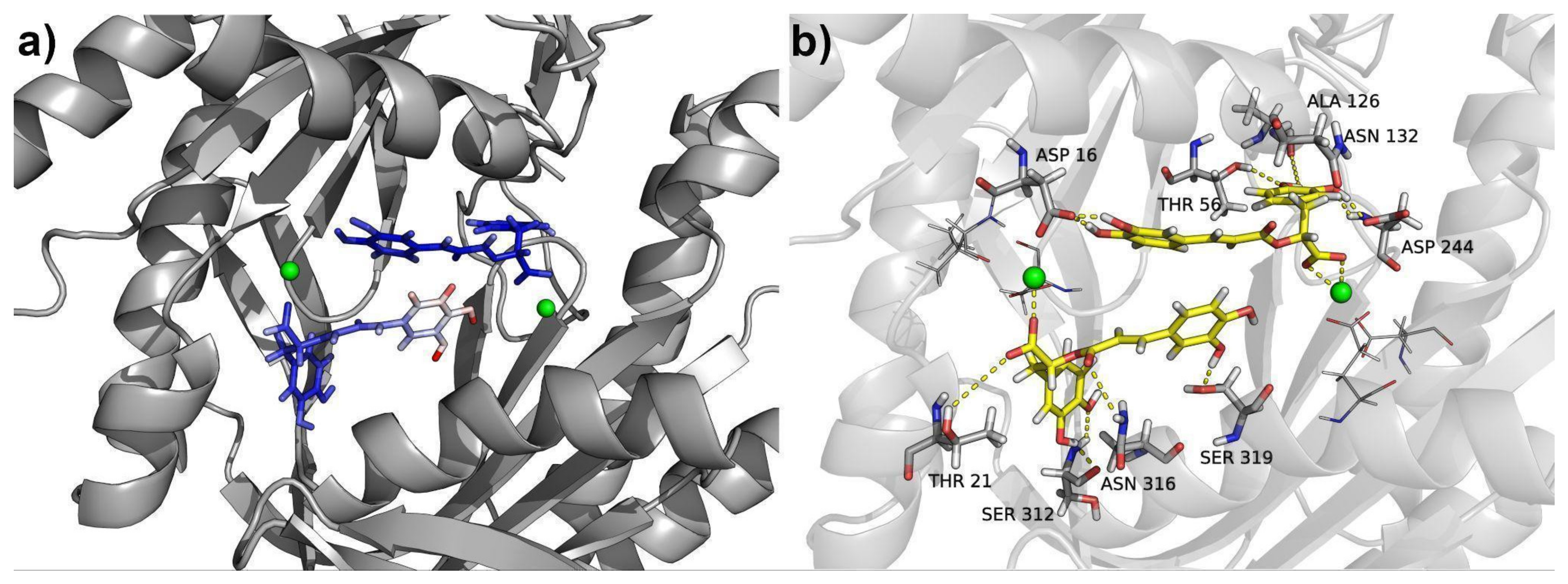
2.3. Molecular Modeling
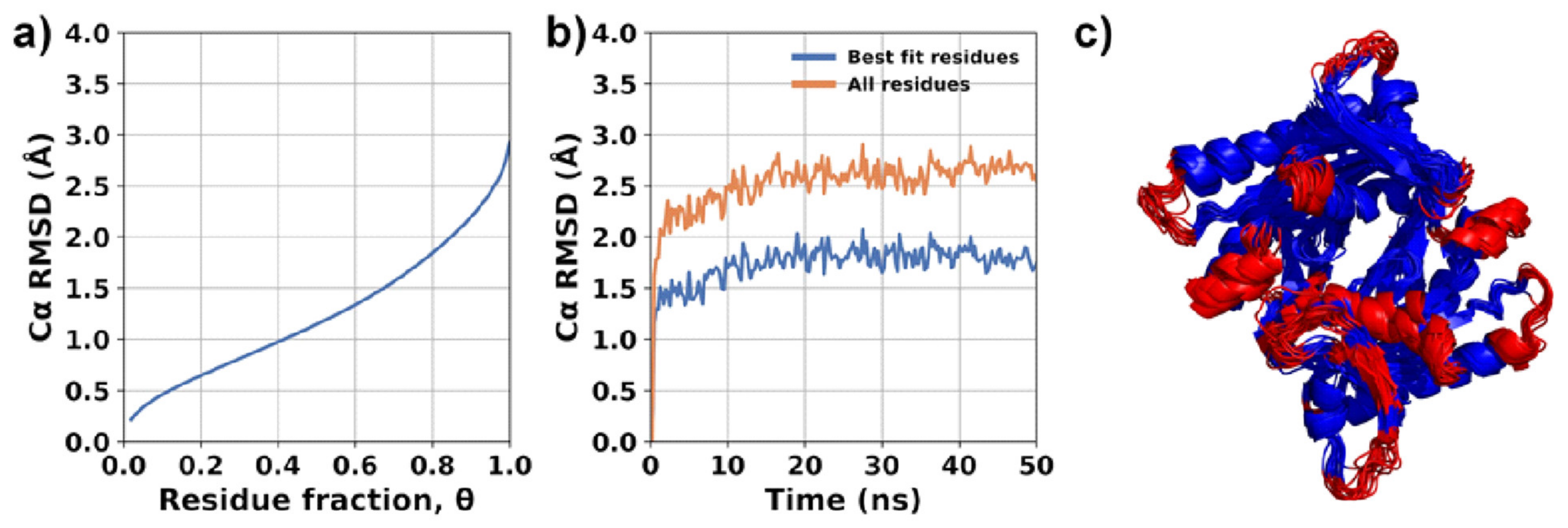
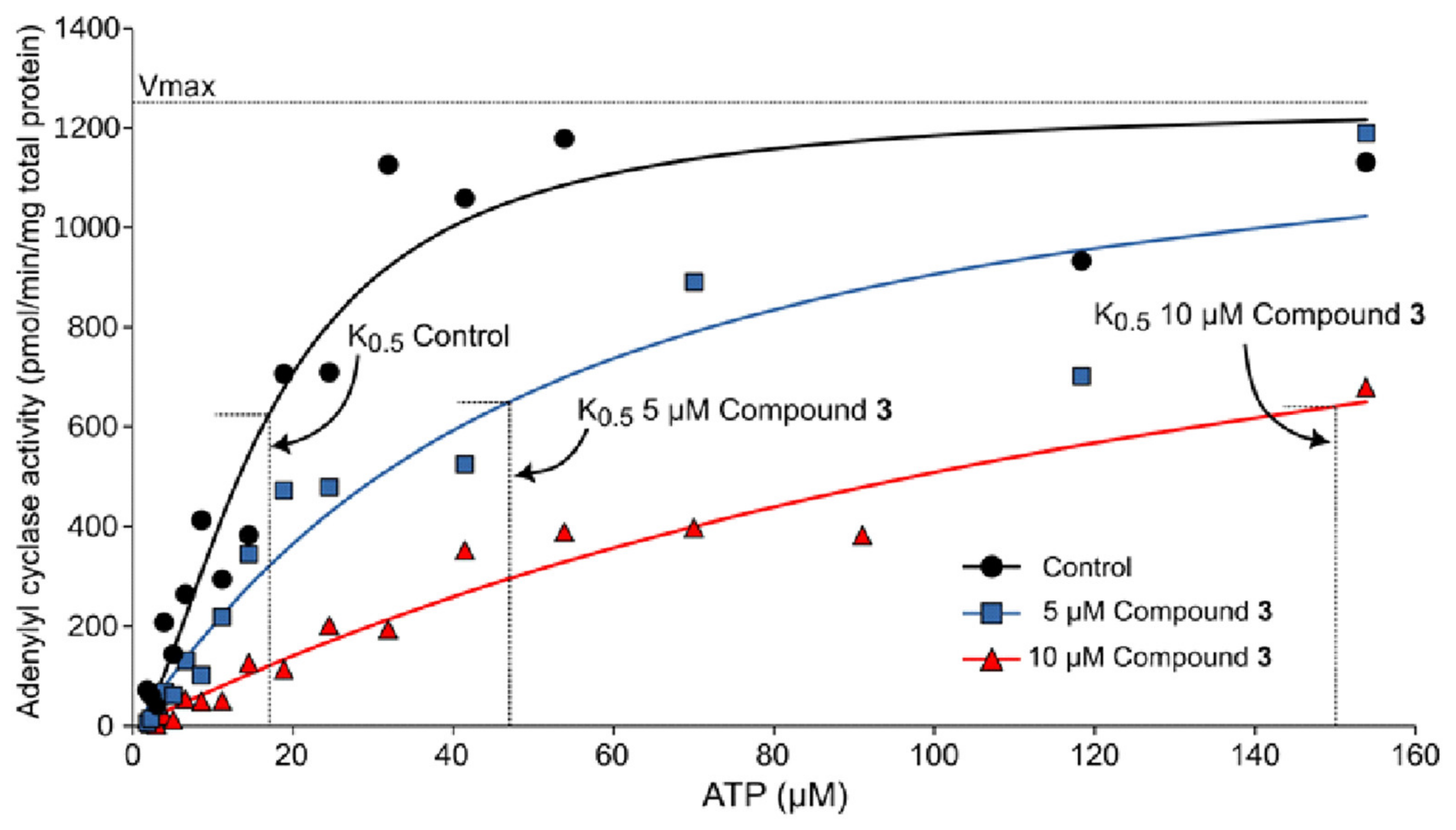
2.4. Experimental Corroboration for the Mechanism of Action of Rosmarinic Acid (3)
3. Materials and Methods
3.1. Materials, Reagents and Equipment
3.2. Plant Material and Extract Preparation
3.3. Quantification of Rosmarinic Acid (3) in Lepechinia Floribunda
3.4. Expression and Enrichment of gNC1 Catalytic Domain
3.5. Adenylyl Cyclase Inhibitory Assays
3.6. cAMP Radiobinding Protein (RBP) Assay
3.7. Determination of the Half-Maximal Inhibitory Concentration (IC50)
3.8. Kinetic Behavior of Rosmarinic Acid (3)
3.9. cAMP-Dependent Protein Kinase (PKA) Purification
3.10. Protein Structure and Energy Minimization
3.11. Docking
3.12. Molecular Dynamics
3.13. Structural Mobility
3.14. Binding Free Energies and Residue Decomposition Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bartelt, L.A.; Platts-Mills, J.A. Giardia: A Pathogen or Commensal for Children in High-Prevalence Settings? Curr. Opin. Infect. Dis. 2016, 29, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Dixon, B.R. Giardia duodenalis in Humans and Animals—Transmission and Disease. Res. Vet. Sci. 2021, 135, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, M.; Halai, K.; Jeffreys, D.; Hunter, P.R. The Prevalence of Giardia Infection in Dogs and Cats, a Systematic Review and Meta-Analysis of Prevalence Studies from Stool Samples. Vet. Parasitol. 2015, 207, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Zahedi, A. Molecular Epidemiology of Giardiasis from a Veterinary Perspective. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 106, pp. 209–254. ISBN 978-0-12-817720-4. [Google Scholar]
- Certad, G.; Viscogliosi, E.; Chabé, M.; Cacciò, S.M. Pathogenic Mechanisms of Cryptosporidium and Giardia. Trends Parasitol. 2017, 33, 561–576. [Google Scholar] [CrossRef]
- Feng, Y.; Xiao, L. Zoonotic Potential and Molecular Epidemiology of Giardia Species and Giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.C.; Leung, A.A.M.; Wong, A.H.C.; Sergi, C.M.; Kam, J.K.M. Giardiasis: An Overview. Recent Pat. Inflamm. Allergy Drug Discov. 2019, 13, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Riches, A.; Hart, C.J.S.; Trenholme, K.R.; Skinner-Adams, T.S. Anti- Giardia Drug Discovery: Current Status and Gut Feelings. J. Med. Chem. 2020, 63, 13330–13354. [Google Scholar] [CrossRef] [PubMed]
- Krakovka, S.; Ribacke, U.; Miyamoto, Y.; Eckmann, L.; Svärd, S. Characterization of Metronidazole-Resistant Giardia intestinalis Lines by Comparative Transcriptomics and Proteomics. Front. Microbiol. 2022, 13, 834008. [Google Scholar] [CrossRef]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural Products for Human Health: An Historical Overview of the Drug Discovery Approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef]
- Chabán, M.F.; Karagianni, C.; Joray, M.B.; Toumpa, D.; Sola, C.; Crespo, M.I.; Palacios, S.M.; Athanassopoulos, C.M.; Carpinella, M.C. Antibacterial Effects of Extracts Obtained from Plants of Argentina: Bioguided Isolation of Compounds from the Anti-Infectious Medicinal Plant Lepechinia meyenii. J. Ethnopharmacol. 2019, 239, 111930. [Google Scholar] [CrossRef]
- Yeshi, K.; Crayn, D.; Ritmejerytė, E.; Wangchuk, P. Plant Secondary Metabolites Produced in Response to Abiotic Stresses Has Potential Application in Pharmaceutical Product Development. Molecules 2022, 27, 313. [Google Scholar] [CrossRef] [PubMed]
- Kamaraj, C.; Ragavendran, C.; Kumar, R.C.S.; Ali, A.; Khan, S.U.; Mashwani, Z.u.-R.; Luna-Arias, J.P.; Pedroza, J.P.R. Antiparasitic Potential of Asteraceae Plants: A Comprehensive Review on Therapeutic and Mechanistic Aspects for Biocompatible Drug Discovery. Phytomed. Plus 2022, 2, 100377. [Google Scholar] [CrossRef]
- García-Huertas, P.; Cardona-Castro, N. Advances in the Treatment of Chagas Disease: Promising New Drugs, Plants and Targets. Biomed. Pharmacother. 2021, 142, 112020. [Google Scholar] [CrossRef] [PubMed]
- Raoof, G.F.A.; Mohamed, A.W.L. Exploring the Antiparasitic Activity of Medicinal Plants. Univers. J. Pharm. Res. 2020, 5, 67–71. [Google Scholar] [CrossRef]
- Selener, M.G.; Elso, O.; Grosso, C.; Borgo, J.; Clavin, M.; Malchiodi, E.L.; Cazorla, S.I.; Redko, F.; Sülsen, V.P. Anti-Trypanosoma cruzi Activity of Extracts from Argentinian Asteraceae Species. Iran. J. Pharm. Res. 2019, 18, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Sülsen, V.; Güida, C.; Coussio, J.; Paveto, C.; Muschietti, L.; Martino, V. In Vitro Evaluation of Trypanocidal Activity in Plants Used in Argentine Traditional Medicine. Parasitol. Res. 2006, 98, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Sülsen, V.P.; Cazorla, S.I.; Frank, F.M.; Redko, F.C.; Anesini, C.A.; Coussio, J.D.; Malchiodi, E.L.; Martino, V.S.; Muschietti, L.V. Trypanocidal and Leishmanicidal Activities of Flavonoids from Argentine Medicinal Plants. Am. J. Trop. Med. Hyg. 2007, 77, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.-L.; Litaudon, M.; Touboul, D.; Queiroz, E.F. Innovative Omics-Based Approaches for Prioritisation and Targeted Isolation of Natural Products—New Strategies for Drug Discovery. Nat. Prod. Rep. 2019, 36, 855–868. [Google Scholar] [CrossRef]
- Fitzgerald, M.; Heinrich, M.; Booker, A. Medicinal Plant Analysis: A Historical and Regional Discussion of Emergent Complex Techniques. Front. Pharmacol. 2020, 10, 1480. [Google Scholar] [CrossRef]
- Davison, E.K.; Brimble, M.A. Natural Product Derived Privileged Scaffolds in Drug Discovery. Curr. Opin. Chem. Biol. 2019, 52, 1–8. [Google Scholar] [CrossRef]
- Sharma, B.; Yadav, D.K. Metabolomics and Network Pharmacology in the Exploration of the Multi-Targeted Therapeutic Approach of Traditional Medicinal Plants. Plants 2022, 11, 3243. [Google Scholar] [CrossRef]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Barboza, G.J.; Cantero, J.E.; Núñez, C.; Pacciaroni, A.; Ariza Espinar, L. Medicinal Plants: A General Review and a Phytochemical and Ethnopharmacological Screening of the Native Argentine Flora. Kurtziana 2009, 34, 7–365. [Google Scholar]
- González, M.L.; Joray, M.B.; Laiolo, J.; Crespo, M.I.; Palacios, S.M.; Ruiz, G.M.; Carpinella, M.C. Cytotoxic Activity of Extracts from Plants of Central Argentina on Sensitive and Multidrug-Resistant Leukemia Cells: Isolation of an Active Principle from Gaillardia megapotamica. Evid. Based Complement. Altern. Med. 2018, 2018, 9185935. [Google Scholar] [CrossRef] [PubMed]
- González, M.L.; Vera, D.M.A.; Laiolo, J.; Joray, M.B.; Maccioni, M.; Palacios, S.M.; Molina, G.; Lanza, P.A.; Gancedo, S.; Rumjanek, V.; et al. Mechanism Underlying the Reversal of Drug Resistance in P-Glycoprotein-Expressing Leukemia Cells by Pinoresinol and the Study of a Derivative. Front. Pharmacol. 2017, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Zuloaga, F.O.; Morrone, O.; Belgrano, M.J. (Eds.) Catálogo de Plantas Vasculares del Cono Sur (Argentina, Sur de Brasil, Chile, Paraguay y Uruguay); Missouri Botanical Garden: Saint Louis, MI, USA, 2008; Volume 1–3, ISBN 978-1-930723-70-2. [Google Scholar]
- Saraullo, V.; Di Siervi, N.; Jerez, B.; Davio, C.; Zurita, A. Synthesis and Degradation of cAMP in Giardia lamblia: Possible Role and Characterization of a Nucleotidyl Cyclase with a Single Cyclase Homology Domain. Biochem. J. 2017, 474, 4001–4017. [Google Scholar] [CrossRef] [PubMed]
- Omori, K.; Kotera, J. Overview of PDEs and Their Regulation. Circ. Res. 2007, 100, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Shemarova, I.V. cAMP-Dependent Signal Pathways in Unicellular Eukaryotes. Crit. Rev. Microbiol. 2009, 35, 23–42. [Google Scholar] [CrossRef]
- Lee, K. Epac: New Emerging cAMP-Binding Protein. BMB Rep. 2021, 54, 149–156. [Google Scholar] [CrossRef]
- Li, Y.; Dillon, T.J.; Takahashi, M.; Earley, K.T.; Stork, P.J.S. Protein Kinase A-Independent Ras Protein Activation Cooperates with Rap1 Protein to Mediate Activation of the Extracellular Signal-Regulated Kinases (ERK) by cAMP. J. Biol. Chem. 2016, 291, 21584–21595. [Google Scholar] [CrossRef]
- Sadeghian, F.; Castaneda, P.G.; Amin, M.R.; Cram, E.J. Functional Insights into Protein Kinase A (PKA) Signaling from C. elegans. Life 2022, 12, 1878. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.S.; Davids, B.J.; Robles, L.D.; Loflin, C.E.; Gillin, F.D.; Chakrabarti, R. Possible Roles of Protein Kinase A in Cell Motility and Excystation of the Early Diverging Eukaryote Giardia lamblia. J. Biol. Chem. 2001, 276, 10320–10329. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.; Schanen, B.; Chakrabarti, D.; Chakrabarti, R. Functional Characterisation of the Regulatory Subunit of Cyclic AMP-Dependent Protein Kinase A Homologue of Giardia lamblia: Differential Expression of the Regulatory and Catalytic Subunits during Encystation. Int. J. Parasitol. 2006, 36, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Vega Hissi, E.G.; De Costa Guardamagna, A.B.; Garro, A.D.; Falcon, C.R.; Anderluh, M.; Tomašič, T.; Kikelj, D.; Yaneff, A.; Davio, C.A.; Enriz, R.D.; et al. A Potent N -(Piperidin-4-yl)-1 H -pyrrole-2-carboxamide Inhibitor of Adenylyl Cyclase of G. lamblia: Biological Evaluation and Molecular Modelling Studies. ChemMedChem 2021, 16, 2094–2105. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.C.; Sprang, S.R. Structures, Mechanism, Regulation and Evolution of Class III Nucleotidyl Cyclases. In Reviews of Physiology Biochemistry and Pharmacology; Amara, S.G., Bamberg, E., Gudermann, T., Hebert, S.C., Jahn, R., Lederer, W.J., Lill, R., Miyajima, A., Offermanns, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 105–140. ISBN 978-3-540-39688-8. [Google Scholar]
- Jaleel, M.; Shenoy, A.R.; Visweswariah, S.S. Tyrphostins Are Inhibitors of Guanylyl and Adenylyl Cyclases. Biochemistry 2004, 43, 8247–8255. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Espiritu, L.; Kleinboelting, S.; Navarrete, F.A.; Alvau, A.; Visconti, P.E.; Valsecchi, F.; Starkov, A.; Manfredi, G.; Buck, H.; Adura, C.; et al. Discovery of LRE1 as a Specific and Allosteric Inhibitor of Soluble Adenylyl Cyclase. Nat. Chem. Biol. 2016, 12, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Saalau-Bethell, S.M.; Berdini, V.; Cleasby, A.; Congreve, M.; Coyle, J.E.; Lock, V.; Murray, C.W.; O’Brien, M.A.; Rich, S.J.; Sambrook, T.; et al. Crystal Structure of Human Soluble Adenylate Cyclase Reveals a Distinct, Highly Flexible Allosteric Bicarbonate Binding Pocket. ChemMedChem 2014, 9, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Kleinboelting, S.; Ramos-Espiritu, L.; Buck, H.; Colis, L.; Van Den Heuvel, J.; Glickman, J.F.; Levin, L.R.; Buck, J.; Steegborn, C. Bithionol Potently Inhibits Human Soluble Adenylyl Cyclase through Binding to the Allosteric Activator Site. J. Biol. Chem. 2016, 291, 9776–9784. [Google Scholar] [CrossRef]
- Crespo, M.I.; Chabán, M.F.; Lanza, P.A.; Joray, M.B.; Palacios, S.M.; Vera, D.M.A.; Carpinella, M.C. Inhibitory Effects of Compounds Isolated from Lepechinia meyenii on Tyrosinase. Food Chem. Toxicol. 2019, 125, 383–391. [Google Scholar] [CrossRef]
- Guan, H.; Luo, W.; Bao, B.; Cao, Y.; Cheng, F.; Yu, S.; Fan, Q.; Zhang, L.; Wu, Q.; Shan, M. A Comprehensive Review of Rosmarinic Acid: From Phytochemistry to Pharmacology and Its New Insight. Molecules 2022, 27, 3292. [Google Scholar] [CrossRef]
- Petersen, M. Rosmarinic Acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef]
- Marchev, A.S.; Vasileva, L.V.; Amirova, K.M.; Savova, M.S.; Koycheva, I.K.; Balcheva-Sivenova, Z.P.; Vasileva, S.M.; Georgiev, M.I. Rosmarinic Acid—From Bench to Valuable Applications in Food Industry. Trends Food Sci. Technol. 2021, 117, 182–193. [Google Scholar] [CrossRef]
- Bouammali, H.; Zraibi, L.; Ziani, I.; Merzouki, M.; Bourassi, L.; Fraj, E.; Challioui, A.; Azzaoui, K.; Sabbahi, R.; Hammouti, B.; et al. Rosemary as a Potential Source of Natural Antioxidants and Anticancer Agents: A Molecular Docking Study. Plants 2023, 13, 89. [Google Scholar] [CrossRef]
- Almela, L.; Sánchez-Muñoz, B.; Fernández-López, J.A.; Roca, M.J.; Rabe, V. Liquid Chromatograpic–Mass Spectrometric Analysis of Phenolics and Free Radical Scavenging Activity of Rosemary Extract from Different Raw Material. J. Chromatogr. A 2006, 1120, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulou, V.; Paloukopoulou, C.; Karioti, A.; Maloupa, E.; Grigoriadou, K. Rosmarinic Acid Production from Origanum dictamnus L. Root Liquid Cultures In Vitro. Plants 2023, 12, 299. [Google Scholar] [CrossRef] [PubMed]
- Trivellini, A.; Lucchesini, M.; Maggini, R.; Mosadegh, H.; Villamarin, T.S.S.; Vernieri, P.; Mensuali-Sodi, A.; Pardossi, A. Lamiaceae Phenols as Multifaceted Compounds: Bioactivity, Industrial Prospects and Role of “Positive-Stress”. Ind. Crops Prod. 2016, 83, 241–254. [Google Scholar] [CrossRef]
- Sánchez-Campillo, M.; Gabaldon, J.A.; Castillo, J.; Benavente-García, O.; Del Baño, M.J.; Alcaraz, M.; Vicente, V.; Alvarez, N.; Lozano, J.A. Rosmarinic Acid, a Photo-Protective Agent against UV and Other Ionizing Radiations. Food Chem. Toxicol. 2009, 47, 386–392. [Google Scholar] [CrossRef]
- Lešnik, S.; Furlan, V.; Bren, U. Rosemary (Rosmarinus officinalis L.): Extraction Techniques, Analytical Methods and Health-Promoting Biological Effects. Phytochem. Rev. 2021, 20, 1273–1328. [Google Scholar] [CrossRef]
- Case, D.A.; Betz, R.M.; Cerutti, D.S.; Cheatham, T.E.I.; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Homeyer, N.; et al. AMBER 16. 2016. Available online: https://ambermd.org/doc12/Amber16.pdf (accessed on 1 February 2022).
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Meagher, K.L.; Redman, L.T.; Carlson, H.A. Development of Polyphosphate Parameters for Use with the AMBER Force Field. J. Comput. Chem. 2003, 24, 1016–1025. [Google Scholar] [CrossRef] [PubMed]
- Bradbrook, M.G.; Gleichmann, T.; Harrop, S.J.; Habash, J.; Raftery, J.; Kalb, J.; Yariv, J.; Hillier, I.H.; Helliwell, J.R. X-Ray and Molecular Dynamics Studies of Concanavalin-A Glucoside and Mannoside Complexes Relating Structure to Thermodynamics of Binding. J. Chem. Soc. Faraday Trans. 1998, 94, 1603–1611. [Google Scholar] [CrossRef]
- Izaguirre, J.A.; Catarello, D.P.; Wozniak, J.M.; Skeel, R.D. Langevin Stabilization of Molecular Dynamics. J. Chem. Phys. 2001, 114, 2090–2098. [Google Scholar] [CrossRef]
- Martínez, L. Automatic Identification of Mobile and Rigid Substructures in Molecular Dynamics Simulations and Fractional Structural Fluctuation Analysis. PLoS ONE 2015, 10, e0119264. [Google Scholar] [CrossRef] [PubMed]
- Andreani, R.; Martínez, J.M.; Martínez, L.; Yano, F. Continuous Optimization Methods for Structure Alignments. Math. Program. 2007, 112, 93–124. [Google Scholar] [CrossRef]
- Martínez, L.; Andreani, R.; Martínez, J.M. Convergent Algorithms for Protein Structural Alignment. BMC Bioinform. 2007, 8, 306. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Cheng, W.-X.; Li, C.; Pan, X.-L.; Xie, X.-G.; Li, T.-H. DFT Study on the Antioxidant Activity of Rosmarinic Acid. J. Mol. Struct. Theochem. 2005, 719, 177–183. [Google Scholar] [CrossRef]
- Trócsányi, E.; György, Z.; Zámboriné-Németh, É. New Insights into Rosmarinic Acid Biosynthesis Based on Molecular Studies. Curr. Plant Biol. 2020, 23, 100162. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Cryst. 1993, 26, 283–291. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Family | Status a | Common Name | Yield (%) b | Voucher UCCOR Number |
|---|---|---|---|---|---|
| Achyrocline satureioides (Lam.) DC. | Asteraceae | N | marcela hembra | 4.5 | 140 |
| Baccharis coridifolia DC. | Asteraceae | N | mio-mio | 2.2 | 147 |
| Calceolaria parviflora Benth. | Calceolariaceae | E | zapatito | 6.1 | 451 |
| Dimerostemma aspilioides (Griseb.) M.D. Moraes | Asteraceae | E | - | 5.5 | 246 |
| Flourensia oolepis S.F. Blake | Asteraceae | E | chilca | 23.0 | 135 |
| Gaillardia megapotamica (Spreng.) Baker | Asteraceae | E | topasaire | 13.76 | 127 |
| Lepechinia floribunda (Benth.) Epling | Lamiaceae | N | salvia blanca | 3.7 | 195 |
| Lepechinia meyenii (Walp.) Epling | Lamiaceae | N | - | 3.8 | 233 |
| Ligaria cuneifolia (Ruiz & Pav.) Tiegh. (parasitizing Vachellia sp.) | Loranthaceae | N | liga | 4.4 | 518 |
| Lithrea molleoides (Vell.) Engl. | Anacardiaceae | N | molle | 10.9 | 183 |
| Melia azedarach L | Meliaceae | Nat | paraíso | 8.7 | 229 c |
| Prosopis alba Griseb. | Fabaceae | N | algarrobo blanco | 8.5 | 255 |
| Senecio viravira Hieron | Asteraceae | N | viravira | 3.9 | 181 |
| Solanum atriplicifolium Gillies ex Nees | Solanaceae | N | - | 18.08 | 528 |
| Plant Extract | IC50 (µg/mL) a |
|---|---|
| Achyrocline satureioides | 51.7 ± 2.5 |
| Baccharis coridifolia | 35.1 ± 6.4 |
| Lepechinia floribunda | 9.4 ± 0.8 |
| Lepechinia meyenii | 31.3 ± 3.2 |
| Lithrea molleoides | 67.0 ± 4.8 |
| Hydrogen Bond (Acceptor ||| Donor) | Fraction | Average Distance (Å) |
|---|---|---|
| Compound 3(site 1)@O1 ||| THR56@OG1HG1 | 0.6746 | 2.7662 |
| ASP 16@OD2 ||| compound 3(site 1)@O4H13 | 0.6384 | 2.6005 |
| ASP 16@OD2 ||| compound 3(site 1)@O3H12 | 0.6326 | 2.5826 |
| ASP 244@OD1 ||| compound 3(site 1)@O2H11 | 0.5018 | 2.6209 |
| ALA 126@O ||| compound 3(site 1)@O1H10 | 0.3572 | 2.7751 |
| compound 3(site 1)@O6 ||| ASN 132@ND2HD21 | 0.104 | 2.8439 |
| ASP 244@OD2 ||| compound 3(site 1)@O2H11 | 0.0934 | 2.6451 |
| SER 312@O ||| compound 3(site 2)@O2H11 | 0.6446 | 2.8028 |
| Compound 3(site 2) @O3 ||| SER 319@OGHG | 0.2506 | 2.7949 |
| Compound 3(site 2)@O7 ||| THR 21@OG1HG1 | 0.1232 | 2.7127 |
| Compound 3(site 2)@O1 ||| SER 312@NH | 0.1078 | 2.9284 |
| SER 319@OG ||| compound 3(site 2)@O3H12 | 0.0904 | 2.8027 |
| Rosmarinic Acid (3) | K0.5 | Vmax (pmol/min/mg Total Protein) | Hill Coefficient |
|---|---|---|---|
| 0 (DMSO control) | 17.0 ± 1.9 | 1249 ± 48 | 1.64 ± 0.32 |
| 5 µM | 47.0 ± 8.9 | 1300 ± 100 | 1.10 ± 0.08 |
| 10 µM | 150.0 ± 16.9 | 1283 ± 93 | 1.04 ± 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zurita, A.; Vega Hissi, E.; Cianci Romero, A.; Luján, A.M.; Salido, S.; Yaneff, A.; Davio, C.; Cobo, J.; Carpinella, M.C.; Enriz, R.D. Rosmarinic Acid Present in Lepechinia floribunda and Lepechinia meyenii as a Potent Inhibitor of the Adenylyl Cyclase gNC1 from Giardia lamblia. Plants 2024, 13, 646. https://doi.org/10.3390/plants13050646
Zurita A, Vega Hissi E, Cianci Romero A, Luján AM, Salido S, Yaneff A, Davio C, Cobo J, Carpinella MC, Enriz RD. Rosmarinic Acid Present in Lepechinia floribunda and Lepechinia meyenii as a Potent Inhibitor of the Adenylyl Cyclase gNC1 from Giardia lamblia. Plants. 2024; 13(5):646. https://doi.org/10.3390/plants13050646
Chicago/Turabian StyleZurita, Adolfo, Esteban Vega Hissi, Agostina Cianci Romero, Adela María Luján, Sofía Salido, Agustín Yaneff, Carlos Davio, Justo Cobo, María Cecilia Carpinella, and Ricardo Daniel Enriz. 2024. "Rosmarinic Acid Present in Lepechinia floribunda and Lepechinia meyenii as a Potent Inhibitor of the Adenylyl Cyclase gNC1 from Giardia lamblia" Plants 13, no. 5: 646. https://doi.org/10.3390/plants13050646