
Effects of Florivory on Floral Visitors and Reproductive Success of Sagittaria lancifolia (Alismataceae) in a Mexican Wetland

Abstract
:1. Introduction
2. Results
2.1. Florivores and Floral Damage
2.2. Florivory and Floral Visitors
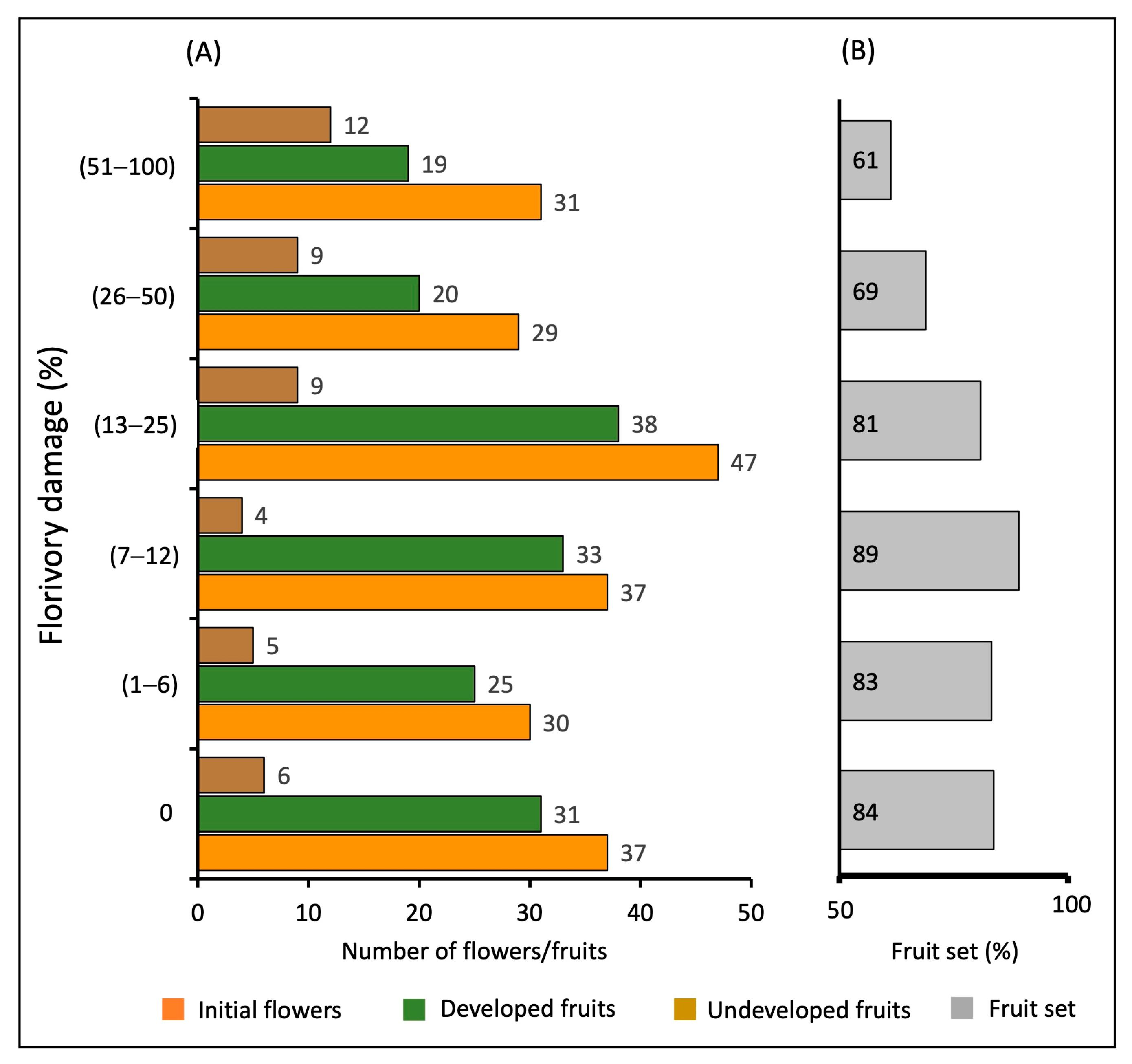
2.3. Florivory and Female Reproductive Success
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. Study Species
4.3. Characterization of Florivory
4.4. Floral Visitors on Damaged Flowers
4.5. Effect of Florivory on Female Reproductive Success
4.6. Evaluation of the Effect of Flower Manipulation on Fruit and Seed Set
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lohman, D.J.; Zangerl, A.R.; Berenbaum, M.R. Impact of floral herbivory by parsnip webworm (Oecophoridae: Depressaria pastinacella Duponchel) on pollination and fitness of wild parsnip (Apiaceae: Pastinaca sativa L.). Amer. Midl. Naturalist 1996, 136, 407–412. [Google Scholar] [CrossRef]
- McCall, A.C.; Irwin, R.E. Florivory: The intersection of pollination and herbivory. Ecol. Lett. 2006, 9, 1351–1365. [Google Scholar] [CrossRef]
- Del Carlo, S. Distribución Espacial de Recursos Florales y Efectos de Antagonistas y Mutualistas en el Éxito Reproductivo de Eriotheca gracilipes (Malvaceae): Una Espécie de Cerrado. Ph.D. Thesis, Instituto de Biociencias de la Universidad de Sao Paulo, Departamento de Ecología, Sao Paulo, Brazil, 13 December 2007. [Google Scholar] [CrossRef]
- Sánchez-Lafuente, A.M. Corolla herbivory, pollination success and fruit predation in complex flowers: An experimental study with Linaria lilacina (Scrophulariaceae). Ann. Bot. 2007, 99, 355–364. [Google Scholar] [CrossRef]
- Xiao, L.; Labandeira, C.; Dilcher, D.; Ren, D. Florivory of Early Cretaceous flowers by functionally diverse insects: Implications for early angiosperm pollination. Proc. R. Soc. B 2021, 288, 20210320. [Google Scholar] [CrossRef] [PubMed]
- Boaventura, M.G.; Villamil, N.; Teixido, A.L.; Tito, R.; Vasconcelos, H.L.; Silveira, F.A.; Cornelissen, T. Revisiting florivory: An integrative review and global patterns of a neglected interaction. New Phytol. 2022, 233, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Mosquera, Y.; Mondragón, D.; Santos-Moreno, A. Vertebrate florivory of vascular epiphytes: The case of a bromeliad. Braz. J. Biol. 2019, 79, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Krupnick, G.A.; Weis, A.E. The effect of floral herbivory on male and female reproductive success in Isomeris arborea. Ecology 1999, 80, 135–149. [Google Scholar] [CrossRef]
- Tsuji, K.; Ohgushi, T. Florivory indirectly decreases the plant reproductive output through changes in pollinator attraction. Ecol. Evol. 2018, 8, 2993–3001. [Google Scholar] [CrossRef] [PubMed]
- Carper, A.L.; Adler, L.S.; Irwin, R.E. Effects of florivory on plant-pollinator interactions: Implications for male and female components of plant reproduction. Am. J. Bot. 2016, 103, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, K.; Dhami, M.K.; Cross, D.J.; Rice, C.P.; Romano, N.H.; Fukami, T. Florivory and pollinator visitation: A cautionary tale. AoB Plants 2016, 8, plw36. [Google Scholar] [CrossRef] [PubMed]
- Glaettli, M.; Barrett, S.C.H. Pollinator responses to variation in floral display and flower size in dioecious Sagittaria latifolia (Alismataceae). New Phytol. 2008, 179, 1193–1201. [Google Scholar] [CrossRef]
- González-Browne, C.; Murúa, M.M.; Navarro, L.; Medel, R. Does plant origin influence the fitness impact of flower damage? A meta-analysis. PLoS ONE 2016, 11, e0146437. [Google Scholar] [CrossRef]
- Soper-Gorden, N.; Adler, L. Florivory shapes both leaf and floral interactions. Ecosphere 2016, 7, e01326. [Google Scholar] [CrossRef]
- Moreira da Silva, G.; Araújo Rodarte, A.T.; Rodrigues Benavides, C.; Alves de Lima, H. Florivory in two Chamaecrista species: Are secretory trichomes effective against florivores? Plant Species Biol. 2022, 37, 339–348. [Google Scholar] [CrossRef]
- Leavitt, H.; Robertson, I.C. Petal herbivory by chrysomelid beetles (Phyllotreta sp.) is detrimental to pollination and seed production in Lepidium papilliferum (Brassicaceae). Ecol. Entomol. 2006, 31, 657–660. [Google Scholar] [CrossRef]
- McCall, A.C. Florivory affects pollinator visitation and female fitness in Nemophila menziesii. Oecologia 2008, 155, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Cascante-Marín, A.; Wolf, J.H.D.; Oostermeijer, J.G.B. Wasp florivory decreases reproductive success in an epiphytic bromeliad. Plant Ecol. 2009, 203, 149–153. [Google Scholar] [CrossRef]
- Sõber, V.; Moora, M.; Teder, T. Florivores decrease pollinator visitation in a self-incompatible plant. Basic Appl. Ecol. 2010, 11, 669–675. [Google Scholar] [CrossRef]
- Ye, Z.-M.; Jin, X.-F.; Wang, Q.-F.; Yang, C.-F.; Inouye, D.W. Pollinators shift to nectar robbers when florivory occurs, with effects on reproductive success in Iris bulleyana (Iridaceae). Plant Biol. 2017, 19, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Ramos, D.; Mandujano, M.C. Florivory effects on pollinator preference and the reproductive output of a threatened living rock cactus, Ariocarpus retusus (Cactaceae). Haseltonia 2019, 25, 133–139. [Google Scholar] [CrossRef]
- Haas, S.M.; Lortie, C.J. A systematic review of the direct and indirect effects of herbivory on plant reproduction mediated by pollination. PeerJ 2020, 8, e9049. [Google Scholar] [CrossRef]
- Vega-Polanco, M.; Rodríguez-Islas, L.A.; Escalona-Domenech, R.Y.; Cruz-López, L.; Rojas, J.C.; Solís-Montero, L. Does florivory affect the attraction of floral visitors to buzz-pollinated Solanum rostratum? Arthropod-Plant Interact. 2020, 14, 41–56. [Google Scholar] [CrossRef]
- Sabino Oliveira, A.C.; Thiabo Souza, J.; Garcia de Brito, V.L.; Almeida, N.M. Attraction of florivores and larcenists and interaction between antagonists in Senna rugosa (Fabaceae). Arthropod-Plant Interact. 2021, 15, 535–544. [Google Scholar] [CrossRef]
- de Matos Costa, K.; Yvila Santos, B.; Messias de Almeida, N.; Melo Santos, A.M.; Buril, M.T.; Leite, A.V. The effects of florivory on floral attractiveness and fruit production in Daustinia montana (Convolvulaceae). Flora 2022, 294, 152122. [Google Scholar] [CrossRef]
- Barrett, S.C. The evolution of plant sexual diversity. Nat. Rev. Genet. 2002, 3, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, M.R.; Lanfear, R.; Mitchell, R.J.; Karron, J.D. Plant mating systems often vary widely among populations. Front. Ecol. Evol. 2018, 6, 38. [Google Scholar] [CrossRef]
- Penet, L.; Collin, C.L.; Ashman, T.-L. Florivory increases selfing: An experimental study in the wild strawberry, Fragaria virginiana. Plant Biol. 2009, 11, 38–45. [Google Scholar] [CrossRef]
- Renner, S.S.; Ricklefs, R.E. Dioecy and its correlates in the flowering plants. Am. J. Bot. 1995, 82, 596–606. [Google Scholar] [CrossRef]
- Cardel, Y.J.; Koptur, S. Effects of florivory on the pollination of flowers: An experimental field study with a perennial plant. Int. J. Plant Sci. 2010, 171, 283–292. [Google Scholar] [CrossRef]
- Hillier, N.K.; Evans, E.; Evans, R.C. Novel insect florivory strategy initiates autogamy in unopened allogamous flowers. Sci. Rep.-UK 2018, 8, 17077. [Google Scholar] [CrossRef]
- Jabbour, F.; Espinosa, F.; Dejonghe, Q.; Le Péchon, T. Development and evolution of unisexual flowers: A review. Plants 2022, 11, 155. [Google Scholar] [CrossRef]
- Teixido, A.L.; Méndez, M.; Valladares, F. Flower size and longevity influence florivory in the large-flowered shrub Cistus ladanifer. Acta Oecol. 2011, 37, 418–421. [Google Scholar] [CrossRef]
- Tsuji, K.; Sota, T. Sexual differences in flower defense and correlated male-biased florivory in a plant–florivore system. Oikos 2010, 119, 1848–1853. [Google Scholar] [CrossRef]
- McCall, A.C.; Barr, C.M. Why do florivores prefer hermaphrodites over females in Nemophila menziesii (Boraginaceae)? Oecologia 2012, 170, 147–157. [Google Scholar] [CrossRef]
- Tsuji, K.; Sota, T. Florivores on the dioecious shrub Eurya japonica and the preferences and performances of two polyphagous geometrid moths on male and female plants. Entomol. Sci. 2013, 16, 291–297. [Google Scholar] [CrossRef]
- Bouchard, P.; Smith, A.B.T.; Douglas, H.; Gimmei, M.L.; Brunke, A.J.; Kanda, K. Biodiversity of Coleoptera. In Insect Biodiversity: Science and Society, 2nd ed.; Foottit, R.G., Adler, P.H., Eds.; John Wiley & Sons Ltd: Hoboken, NJ, USA, 2017; Volume 1. [Google Scholar] [CrossRef]
- Ferreira, C.A.; Torezan-Silingardi, H.M. Implications of the floral herbivory on Malpighiacea plant fitness: Visual aspect of the flower affects the attractiveness to pollinators. Sociobiology 2013, 60, 323–328. [Google Scholar] [CrossRef]
- Ortiz, G.L.; Columbano, Y.; de Melo, M.V.; Boaventura, M.G.; Aoki, C.; Cornelissen, T.; Souza, C.S.; Teixido, A.L. Among-species variation in flower size determines florivory in the largest seasonally flooded tropical wetland. Am. J. Bot. 2023, 110, e16186. [Google Scholar] [CrossRef] [PubMed]
- Hipólito, F.P.N.; Chaves, T.R. Florivory and sex ratio in Annona dioica St. Hil. (Annonaceae) in the Pantanal at Nhecolandia, southwestern Brazil. Acta Bot. Bras. 2006, 20, 405–409. [Google Scholar] [CrossRef]
- Huang, S.-Q. Flower dimorphism and maintenance of andromonoecy in Sagittaria guayanensis spp. lappula (Alismataceae). New Phytol. 2003, 157, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Miguez, M.B.; Amela García, M.T. Breeding system, floral biology and pollinators of Sagittaria montevidensis subsp. Montevidensis Cham. et Schlech. (Alismataceae). Aquat Bot. 2019, 158, 103123. [Google Scholar] [CrossRef]
- Figueroa-Castro, D.M. Efecto de la Herbivoría Floral Sobre el Éxito Reproductivo de Dahlia coccínea (Asteraceae) en el Pedregal de San Ángel. Master’s Thesis, Facultad de Ciencias, UNAM, Mexico City, Mexico, 2001. [Google Scholar]
- Botto-Mahan, C.; Ramírez, P.A.; Ossa, C.G.; Medel, R.; Ojeda-Camacho, M.; González, A.V. Floral herbivory affects female reproductive success and pollinator visitation in the perennial herb Alstroemeria ligtu (Alstroemeriaceae). Int. J. Plant Sci. 2011, 172, 1130–1136. [Google Scholar] [CrossRef]
- Althoff, D.M.; Segraves, K.A.; Pellmyr, O. Community context of an obligate mutualism: Pollinator and florivore effects on Yucca filamentosa. Ecology 2005, 86, 905–913. [Google Scholar] [CrossRef]
- Araujo, A.I.; Freitas, L. Is pollen removal or seed set favored by flower longevity in a hummingbird-pollinated Salvia species? Ann. Bot. 2010, 106, 413–419. [Google Scholar] [CrossRef]
- McCall, A.C. Does dose-dependent petal damage affect pollen limitation in an annual plant? Botany 2010, 88, 601–606. [Google Scholar] [CrossRef]
- Kudoh, H.; Whigham, D.F. The effect of petal size manipulation on pollinator/seed-predator mediated female reproductive success of Hibiscus moscheutos. Oecologia 1998, 117, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Casasola, P.; Cejudo-Espinosa, E.; Capistrán-Barradas, A.; Infante-Mata, D.; López-Rosas, H.; Castillo-Campos, G.; Pale-Pale, J.; Campos-Cascaredo, A. Composición florística, diversidad y ecología de humedales herbáceos emergentes en la planicie costera central de Veracruz, México. Bol. Soc. Bot. Méx. 2010, 87, 29–50. [Google Scholar] [CrossRef]
- Castillo-Campos, G.; Medina, A.M.E. Árboles y Arbustos de la Reserva Natural de La Mancha, Veracruz, 1st ed.; Instituto de Ecología A. C. Xalapa, Ver.: Mexico D.F., Mexico, 2005. [Google Scholar]
- Martínez-Adriano, C.A.; Aguirre-Jaimes, A.; Díaz-Castelazo, C. Floristic survey of flowering plants in a tropical coastal ecosystem in Veracruz, Mexico. Bot. Sci. 2016, 94, 185–197. [Google Scholar] [CrossRef]
- Lopez Rosas, H.; Moreno-Casasola, P.; Mendelssohn, I.A. Effects of experimental disturbances on a tropical freshwater marsh invaded by the African grass Echinochloa pyramidalis. Wetlands 2006, 26, 593–604. [Google Scholar] [CrossRef]
- Dorken, M.E.; Friedman, J.; Barrett, S.C.H. The evolution and maintenance of monoecy and dioecy in Sagittaria latifolia (Alismataceae). Evolution 2002, 56, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Novelo, A. Alismataceae. Fascículo 111. In Flora del Bajío y de Regiones Adyacentes; Rzedowski, J., Calderon de Rzedowski, G., Eds.; Instituto de Ecología, A.C., Conacyt, Conabio: Patzcuaro, Mexico, 2003; pp. 1–19. [Google Scholar] [CrossRef]
- Rodríguez-Morales, D. Efecto de la Florivoría Sobre los Visitantes Florales y el Éxito Reproductivo de Sagittaria lancifolia Media (Alismataceae) en el Humedal de La Mancha. Bachelor’s Thesis, Facultad de Biología, Benemérita Universidad Autónoma de Puebla, Mexico City, Mexico, 2013. [Google Scholar]
- Haynes, R.R. Alismataceae. Fascículo 37. In Flora de Veracruz; Gómez-Pompa, A., Moreno, N.P., Eds.; INIREB: Xalapa, Mexico, 1984; pp. 17–23. Available online: https://libros.inecol.mx/index.php/FV/catalog/view/451/508/2510 (accessed on 20 December 2023).
- Dirzo, R.; Domínguez, C.A. Plant-herbivore interactions in Mesoamerican tropical dry forests. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 304–325. [Google Scholar] [CrossRef]
- Ruiz-Guerra, B.; Nieves-Silva, E.; Guevara, R. The role of leaf traits and bird-mediated insect predation on patterns of herbivory in a semiarid environment in central Mexico. Bot. Sci. 2017, 95, 189–201. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontologist Statistics software package for education and data analysis. Paleontol. Electron. 2011, 4, 9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Florivory Damage Category (%) | Fruit Weight (g) | Seed Weight (g) | Number of Seeds per Fruit |
|---|---|---|---|
| 0 | 0.237 ± 0.019 a | 0.213 ± 0.018 a | 1287.3 ± 107.7 a |
| 1–6 | 0.237 ± 0.023 a | 0.214 ± 0.021 a | 1080.3 ± 106.3 ac |
| 7–12 | 0.188 ± 0.012 b | 0.169 ± 0.011 b | 910.7 ± 60.0 bcd |
| 13–25 | 0.120 ± 0.010 b | 0.107 ± 0.010 b | 859.0 ± 80.8 bc |
| 51–100 | 0.146 ± 0.014 b | 0.132 ± 0.013 b | 791.9 ± 77.9 bc |
| Flower Treatment | Seed Weight (g) | Seed Number per Fruit |
|---|---|---|
| Undamaged | 0.208 ± 0.027 | 1251.7 ± 143.2 |
| Florivory | 0.205 ± 0.031 | 1164.1 ± 166.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Morales, D.; Aguirre-Jaimes, A.; García-Franco, J.G. Effects of Florivory on Floral Visitors and Reproductive Success of Sagittaria lancifolia (Alismataceae) in a Mexican Wetland. Plants 2024, 13, 547. https://doi.org/10.3390/plants13040547
Rodríguez-Morales D, Aguirre-Jaimes A, García-Franco JG. Effects of Florivory on Floral Visitors and Reproductive Success of Sagittaria lancifolia (Alismataceae) in a Mexican Wetland. Plants. 2024; 13(4):547. https://doi.org/10.3390/plants13040547
Chicago/Turabian StyleRodríguez-Morales, Dulce, Armando Aguirre-Jaimes, and José G. García-Franco. 2024. "Effects of Florivory on Floral Visitors and Reproductive Success of Sagittaria lancifolia (Alismataceae) in a Mexican Wetland" Plants 13, no. 4: 547. https://doi.org/10.3390/plants13040547