


Structural and Functional Strategies in Cenchrus Species to Combat Environmental Extremities Imposed by Multiple Abiotic Stresses
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Environmental and Soil Physicochemical Traits
2.2. Proportion of Morpho-Physiological Traits
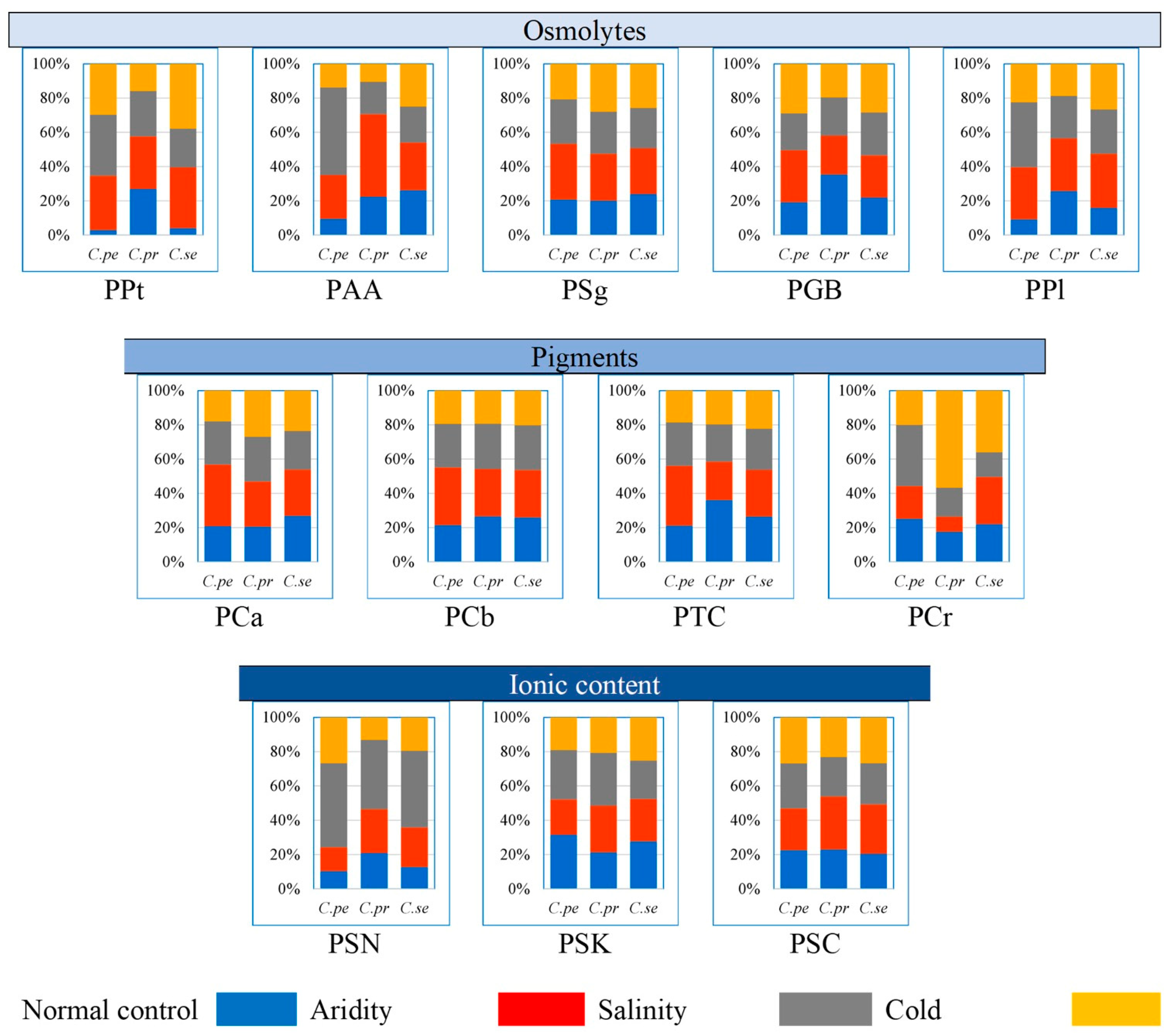
2.3. Proportion of Physiological Traits
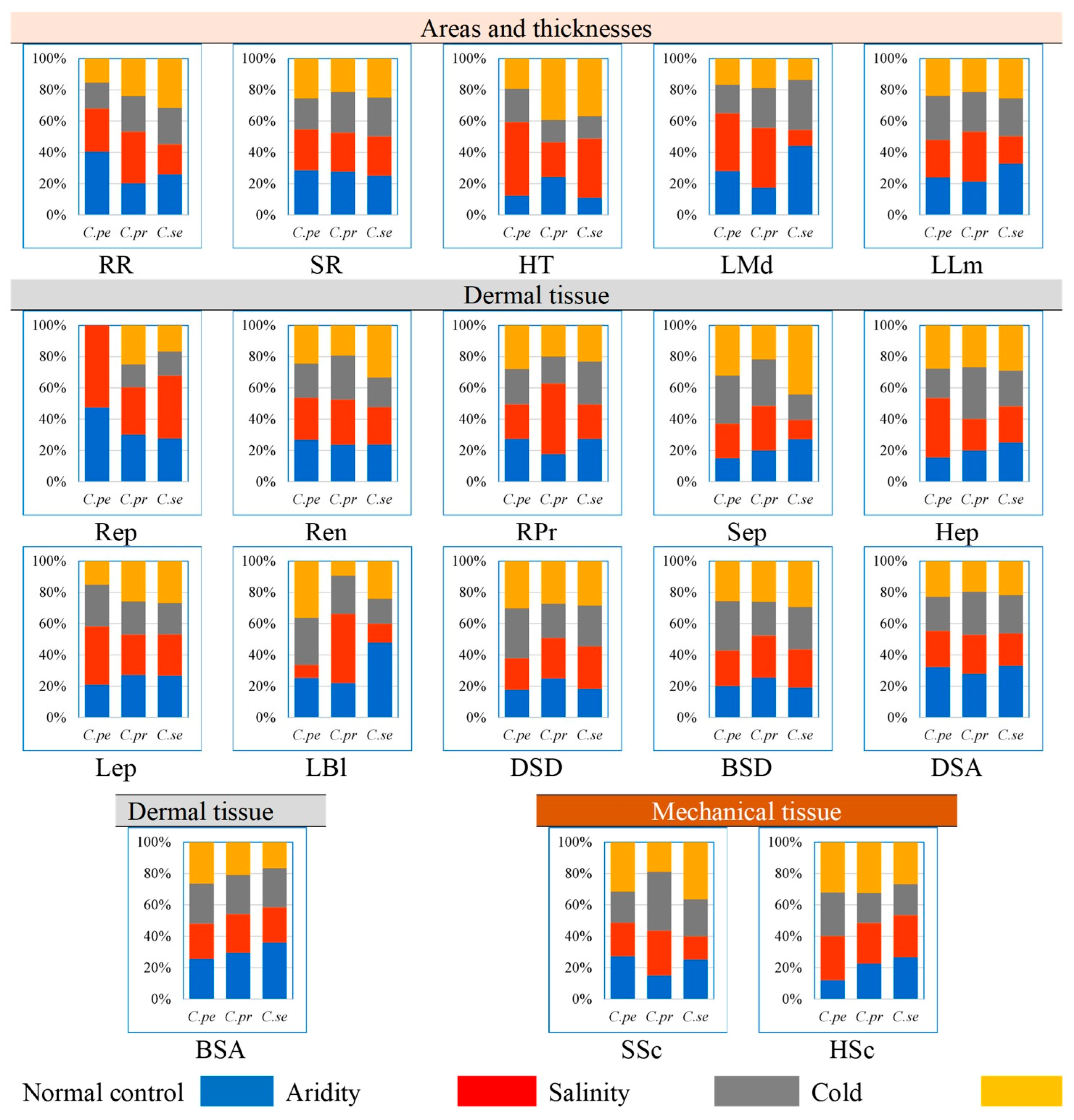
2.4. Proportion of Dermal Tissue
2.5. Proportion of Mechanical Tissue
2.6. Proportion of Areas and Thickness Root, Stem and Leaves
2.7. Proportion of Parenchymatous Tissue
2.8. Proportion of Vascular Tissue
2.9. Pearson’s Correlation Coefficients (p < 0.05) for Environmental/Soil and Morpho-Anatomical and Physiological Traits
2.10. Principal Component Analysis Showing Relationship among Soil/Environmental and Plant Traits
2.11. Relationship between Soil/Environmental and Morpho-Physiological Traits Anatomical Traits
2.12. Relationship between Soil/Environmental and Root/Stem Anatomical Traits
2.13. Relationship between Soil/Environmental and Leaf Sheath/Leaf Anatomical Traits
2.14. Estimated Response of Structural and Functional Traits
3. Discussion
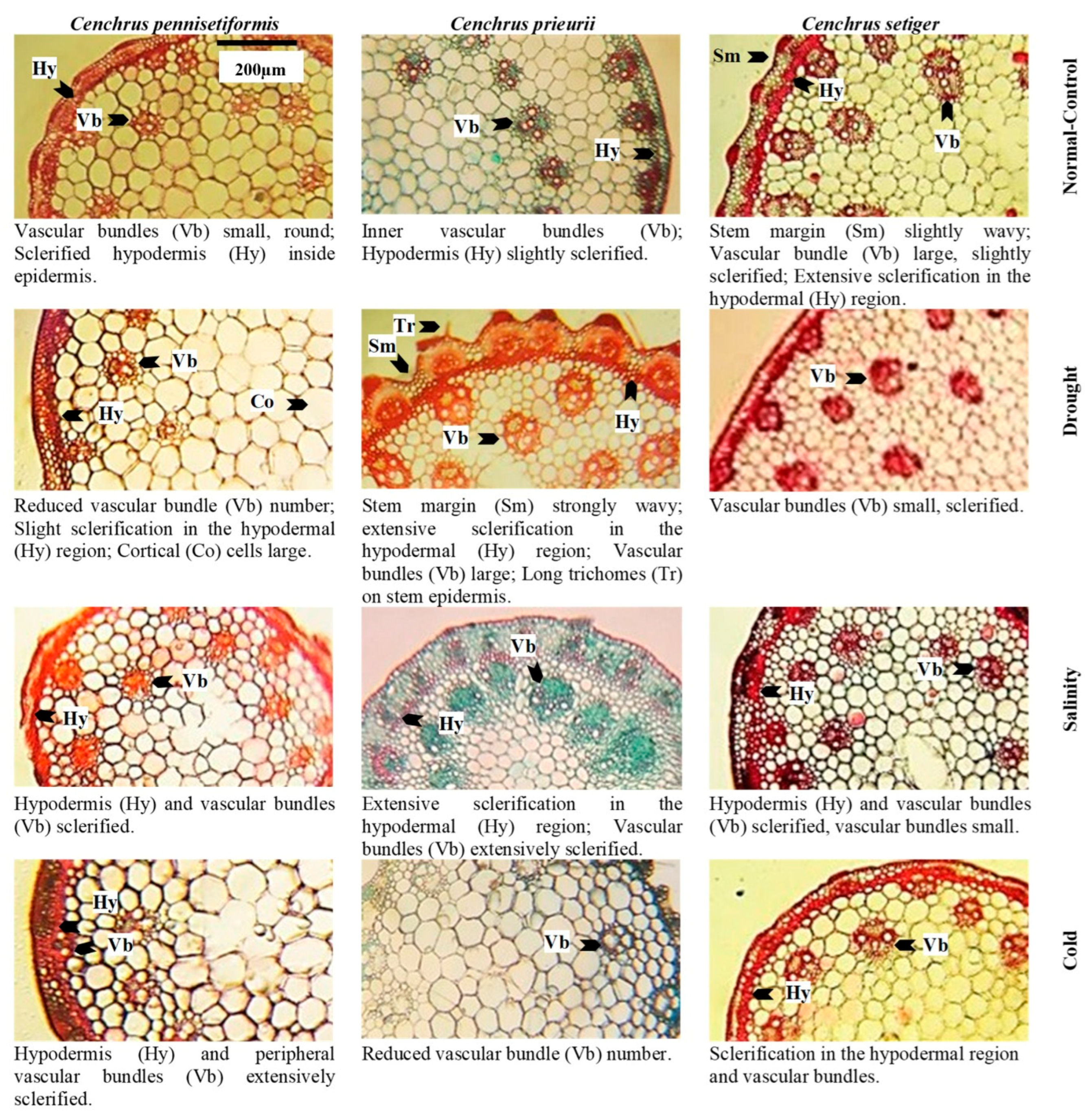
3.1. Cenchrus pennisetiformis
3.2. Cenchrus prieurii
3.3. Cenchrus setiger
4. Materials and Methods
4.1. Sample Collection
4.2. Physiographic Data
4.3. Soil Analysis
4.4. Morphological Traits
4.5. Physiological Traits
4.5.1. Ionic Content
4.5.2. Photosynthetic Pigments
4.5.3. Biochemical Traits
4.6. Anatomical Traits
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parihar, P.; Singh, S.; Singh, R. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Bussotti, F.; Pollastrini, M. Revisiting the concept of stress in forest trees at the time of global change and issues for stress monitoring. Plant Stress 2021, 2, 100013. [Google Scholar] [CrossRef]
- Bashir, S.S.; Hussain, A.; Hussain, S.J.; Wani, O.A.; Zahid Nabi, S.; Dar, N.A.; Mansoor, S. Plant drought stress tolerance: Understanding its physiological, biochemical and molecular mechanisms. Biotechnol. Biotechnol. Equip. 2021, 35, 1912–1925. [Google Scholar] [CrossRef]
- Silva, E.N.; Vieira, S.A.; Ribeiro, R.V.; Ponte, L.F.; Ferreira-Silva, S.L.; Silveira, J.A. Contrasting physiological responses of Jatropha curcas plants to single and combined stresses of salinity and heat. J. Plant Growth Regul. 2013, 32, 159–169. [Google Scholar] [CrossRef]
- Perdomo, J.A.; Conesa, M.À.; Medrano, H.; Ribas-Carbó, M.; Galmés, J. Effects of long-term individual and combined water and temperature stress on the growth of rice, wheat and maize: Relationship with morphological and physiological acclimation. Physiol. Plant 2015, 155, 149–165. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.; Jain, R.; Urwin, P. The Response of Plants to Simultaneous Biotic and Abiotic Stress. In Combined Stresses in Plants; Mahalingam, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 181–201. [Google Scholar]
- Alvarez-Flores, R.; Nguyen-Thi-Truc, A.; Peredo-Parada, S.; Joffre, R.; Winkel, T. Rooting plasticity in wild and cultivated Andean Chenopodium species under soil water deficit. Plant Soil 2018, 425, 479–492. [Google Scholar] [CrossRef]
- Ali, M.; Gul, A.; Hasan, H.; Gul, S.; Fareed, A.; Nadeem, M.; Jamil, M. Cellular mechanisms of drought tolerance in wheat. In Climate Change and Food Security with Emphasis on Wheat; Ozturk, M., Gul, A., Eds.; Academic Press: New York, NY, USA, 2020; pp. 155–167. [Google Scholar]
- Abobatta, W.F. Plant Responses and Tolerance to Extreme Salinity: Learning from Halophyte Tolerance to Extreme Salinity. In Salt and Drought Stress Tolerance in Plants. Signaling and Communication in Plants; Hasanuzzaman, M., Tanveer, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 177–210. [Google Scholar]
- Soliveres, S.; Smit, C.; Maestre, F.T. Moving forward on facilitation research: Response to changing environments and effects on the diversity, functioning and evolution of plant communities. Biol. Rev. 2015, 9, 297–313. [Google Scholar] [CrossRef]
- Ghalambor, C.K.; McKay, J.K.; Carroll, S.P.; Reznick, D.N. Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct. Ecol. 2007, 21, 394–407. [Google Scholar] [CrossRef]
- Fatima, S.; Hameed, M.; Ahmad, F.; Ahmad, M.S.A.; Anwar, M.; Munir, M.; Khalil, S. Dramatic changes in anatomical traits of a C4 grass Chrysopogon serrulatus Trin. (Poaceae) over a 1000 m elevational gradient. J. Mount. Sci. 2023, 20, 1316–1335. [Google Scholar] [CrossRef]
- Bucur, V. Traditional and new materials for the reeds of woodwind musical instruments. Wood Sci. Technol. 2019, 53, 1157–1187. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Babar, M.A. Role of sugars, amino acids and organic acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Merrium, S.; Ali, Z.; Tahir, M.H.N.; Habib-ur-Rahman, M.; Hakeem, S. Leaf rolling dynamics for atmospheric moisture harvesting in wheat plant as an adaptation to arid environments. Environ. Sci. Pollut. Res. 2022, 29, 48995–49006. [Google Scholar] [CrossRef] [PubMed]
- Perlikowski, D.; Augustyniak, A.; Masajada, K.; Skirycz, A.; Soja, A.M.; Michaelis, Ä.; Wolter, G.; Kosmala, A. Structural and metabolic alterations in root systems under limited water conditions in forage grasses of Lolium-Festuca complex. Plant Sci. 2019, 283, 211–223. [Google Scholar] [CrossRef]
- Fukao, T.; Xiong, L. Genetic mechanisms conferring adaptation to submergence and drought in rice: Simple or complex? Curr. Opin. Plant Biol. 2013, 16, 196–204. [Google Scholar] [CrossRef]
- Habbib, H.; Hirel, B.; Spicher, F.; Dubois, F.; Tétu, T. In winter wheat (Triticum aestivum L.), no-till improves photosynthetic nitrogen and water-use efficiency. J. Crop Sci. Biotechnol. 2020, 23, 39–46. [Google Scholar] [CrossRef]
- Feng, Z.; Sun, Q.; Deng, Y.; Sun, S.; Zhang, J.; Wang, B. Study on pathway and characteristics of ion secretion of salt glands of Limonium bicolor. Acta Physiol. Plant 2014, 36, 2729–2741. [Google Scholar] [CrossRef]
- Khurshid, S.; Shoaib, A.; Javaid, A.; Akhtar, F.; Shafiq, M.; Qaisar, U. Management of Fusarium wilt of tomato by soil amendment with Cenchrus pennisetiformis under chromium stress. Physiol. Mol. Plant Pathol. 2017, 97, 58–68. [Google Scholar] [CrossRef]
- Shiran, K.; Noor Mohamed, M.B.; Keerthika, A.; Pareek, K.; Pandey, C.B. Agroforestry systems for arid ecologies in India. Agroforestry for degraded landscapes. Recent. Adv. Emerg. Chall. 2020, 1, 169–188. [Google Scholar]
- Abdi, S.; Dwivedi, A.; Shashi; Kumar, S.; Bhat, V. Development of EST-SSR markers in Cenchrus ciliaris and their applicability in studying the genetic diversity and cross-species transferability. J. Genet. 2019, 98, 101. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.I.; Akhtar, N.; Qureshi, A.S.; Gallacher, D. Perennial forage grass production on the marginal Arabian Peninsula land. Sust. Agric. Rev. 2021, 52, 279–308. [Google Scholar]
- Kalamani, A.; Babu, C.; Iyanar, K.; Sudhagar, R.; Sivakumar, S.D.; Ezhilarasi, T.; Geetha, S. A new high yielding black kolukattai grass variety CO2 (Cenchrus setigerus) suitable for pasture lands. Elect. J. Plant Breed. 2021, 12, 1085–1090. [Google Scholar]
- Halli, H.M.; Govindasamy, P.; Choudhary, M.; Srinivasan, R.; Prasad, M.; Wasnik, V.K.; Pathak, H. Range grasses to improve soil properties, carbon sustainability, and fodder security in degraded lands of semi-arid regions. Sci. Total Environ. 2022, 851, 158211. [Google Scholar] [CrossRef] [PubMed]
- Basharat, S.; Ahmad, F.; Hameed, M.; Naeem, M.S. Defense through foliar anatomical alterations in large spike buffel grass [Cenchrus prieurii (Kunth) Maire] against multiple environmental stresses. Pak. J. Bot. 2023, 55, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Linder, H.P.; Lehmann, C.E.; Archibald, S.; Osborne, C.P.; Richardson, D.M. Global grass (Poaceae) success underpinned by traits facilitating colonization, persistence and habitat transformation. Biol. Rev. 2018, 93, 1125–1144. [Google Scholar] [CrossRef]
- Harun, N.; Shaheen, S.; Ahmad, M.; Bibi, F.; Fatima, K.; Ramzan, S.; Bashir, H. Light and scanning electron microscopic imaging of leaf transverse sections of indigenous fodder grasses of central Punjab, Pakistan. Microsc. Res. Tech. 2022, 85, 2497–2513. [Google Scholar] [CrossRef]
- Ahlem, A.; Lobna, M.; Mohamed, C. Ecophysiological responses of different ploidy levels (tetraploid and hexaploid), of Cenchrus ciliaris to water deficiency conditions. Pak. J. Bot. 2021, 53, 1997–2002. [Google Scholar] [CrossRef]
- Kumar, S.; Saxena, S.; Rai, A.; Radhakrishna, A.; Kaushal, P. Ecological, genetic, and reproductive features of Cenchrus species indicate evolutionary superiority of apomixis under environmental stresses. Ecol. Indic. 2019, 105, 126–136. [Google Scholar] [CrossRef]
- Rafay, M.; Abid, M.; Abdullah, M.; Malik, Z.; Makki, H.A.; Ahmed, Z.; Riaz, M.U. Growth parameters and antioxidant enzymes activities in selected halophytic grass species from Cholistan rangeland, Pakistan under salinity stress. Pak. J. Bot. 2021, 53, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Mann, A.; Lata, C.; Kumar, A. Physiological adaptability of Salvadora oleoides to sodicity and salinity stress. Indian J. Agric. Sci. 2022, 92, 1480–1483. [Google Scholar] [CrossRef]
- Sultan, I.; Khan, I.; Chattha, M.U.; Hassan, M.U.; Barbanti, L.; Calone, R.; Ali, M.; Majeed, S.; Ghani, M.A.; Batool, M.; et al. Improved salinity tolerance in early growth stage of maize through salicylic acid foliar application. Ital. J. Agron. 2021, 16, 1–11. [Google Scholar]
- Ejaz, S.; Hussain, S.; Anjum, M.A.; Ahmad, S. Application of osmolytes in improving abiotic stress tolerance in plant. In Plant Tolerance to Environmental Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 47–58. [Google Scholar]
- Masouleh, S.S.S.; Aldine, N.J.; Sassine, Y.N. The role of organic solutes in the osmotic adjustment of chilling-stressed plants (vegetable, ornamental and crop plants). Ornam. Hortic. 2020, 25, 434–442. [Google Scholar] [CrossRef]
- Soukup, A.; Tylová, E. Apoplastic barriers: Their structure and function from a historical perspective. In Concepts in Cell Biology—History and Evolution; Springer: Berlin/Heidelberg, Germany, 2018; pp. 155–183. [Google Scholar]
- Parvez, I.; Ahmad, I.; Hameed, M.; Ahmad, M.S.A. Plasticity in anatomical traits of Cyperus niveus Retz. along elevational gradient of dry sub-Himalayan mountains. Arid. Land. Res. Manag. 2023, 37, 197–221. [Google Scholar] [CrossRef]
- Ahmad, K.S.; Wazarat, A.; Mehmood, A.; Ahmad, M.S.A.; Tahir, M.M.; Nawaz, F.; Ulfat, A. Adaptations in Imperata cylindrica (L.) Raeusch. and Cenchrus ciliaris L. for altitude tolerance. Biologia 2020, 75, 183–198. [Google Scholar]
- Mata, A.T.; Jorge, T.F.; Pires, M.V.; Antonio, C. Drought stress tolerance in plants: Insights from metabolomics. Drought Stress Toler. Plants Mol. Genet. Perspect. 2016, 2, 187–216. [Google Scholar]
- Wasim, M.A.; Naz, N.; Zehra, S.S. Anatomical characteristic, ionic contents and nutritional potential of buffel grass (Cenchrus ciliaris L.) under high salinity. S. Afr. J. Bot. 2022, 144, 471–479. [Google Scholar] [CrossRef]
- Arafa, A.A.; Khafagy, M.A.; El-Banna, M.F. The effect of glycinebetaine or ascorbic acid on grain germination and leaf structure of sorghum plants grown under salinity stress. Aust. J. Crop Sci. 2009, 3, 294–304. [Google Scholar]
- Wasaya, A.; Zhang, X.; Fang, Q.; Yan, Z. Root phenotyping for drought tolerance: A review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef]
- Ghafar, M.A.; Akram, N.A.; Saleem, M.H.; Wang, J.; Wijaya, L.; Alyemeni, M.N. Ecotypic morphological and physio-biochemical responses of two differentially adapted forage grasses, Cenchrus ciliaris L. and Cyperus arenarius Retz. to drought stress. Sustainability 2021, 13, 8069. [Google Scholar] [CrossRef]
- Nadeem, M.; Anwar-ul-Haq, M.; Saqib, M.; Maqsood, M.; He, Z. Ameliorative effect of silicic acid and silicates on oxidative, osmotic stress, and specific ion toxicity in spring wheat (Triticum aestivum L.) genotypes. J. Soil Sci. Plant Nutr. 2022, 22, 2334–2345. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Morpho-anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. [Google Scholar]
- Matschi, S.; Vasquez, M.F.; Bourgault, R.; Steinbach, P.; Chamness, J.; Kaczmar, N.; Gore, M.A.; Molina, I.; Smith, L.G. Structure-function analysis of the maize bulliform cell cuticle and its potential role in dehydration and leaf rolling. Plant Direct 2020, 4, e00282. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Ni, X.; Chen, C.; Zhang, S.; Chen, X.; Yang, Z.; Li, C. Effects of different water conditions on the biomass, root morphology and aerenchyma formation in bermudagrass (Cynodon dactylon (L.) Pers). BMC Plant Biol. 2022, 22, 266. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.K.; Malaviya, D.R.; Kaushal, P. Genetic improvement of dominant tropical Indian range grasses. Range Manag. Agrofor. 2019, 40, 1–25. [Google Scholar]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar]
- Fatima, S.; Hameed, M.; Ahmad, F.; Ahmad, M.S.A.; Khalil, S.; Munir, M.; Ashraf, M.; Ahmad, I.; Ahmad, K.S.; Kaleem, M. Structural and functional responses in widespread distribution of some dominant grasses along climatic elevation gradients. Flora 2022, 289, 152034. [Google Scholar] [CrossRef]
- Olsen, J.T.; Caudle, K.L.; Johnson, L.C.; Baer, S.G.; Maricle, B.R. Environmental and genetic variation in leaf anatomy among populations of Andropogon gerardii (Poaceae) along a precipitation gradient. Am. J. Bot. 2013, 100, 1957–1968. [Google Scholar] [CrossRef]
- da Silva Lobato, S.M.; dos Santos, L.R.; da Silva, B.R.S.; Paniz, F.P.; Batista, B.L.; da Silva Lobato, A.K. Root-differential modulation enhances nutritional status and leaf anatomy in pigeonpea plants under water deficit. Flora 2020, 262, 151519. [Google Scholar] [CrossRef]
- Chen, I.; Li, K.T.; Tsang, C.H. Silicified bulliform cells of Poaceae: Morphological characteristics that distinguish subfamilies. Bot. Stud. 2020, 61, 1–25. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of plant responses to water stress and related genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Kasirajan, L.; Valiyaparambth, R.; Kamaraj, K.; Sebastiar, S.; Hoang, N.V.; Athiappan, S.; Subramanian, K. Deep sequencing of suppression subtractive library identifies differentially expressed transcripts of Saccharum spontaneum exposed to salinity stress. Physiol. Plant. 2022, 174, e13645. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Ahmar, S.; Ali, B.; Saleem, M.H.; Khan, M.U.; Zhou, W.; Liu, S. The role of membrane transporters in plant growth and development, and abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 12792. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Vilagrosa, A.; Alonso-Forn, D.; Ferrio, J.P.; Sancho-Knapik, D.; Gil-Pelegrín, E. Living in drylands: Functional adaptations of trees and shrubs to cope with high temperatures and water scarcity. Forests 2020, 11, 1028. [Google Scholar] [CrossRef]
- Veldhuis, E.R.; Schrama, M.; Staal, M.; Elzenga, J.T.M. Plant stress-tolerance traits predict salt marsh vegetation patterning. Front. Mar. Sci. 2019, 5, 501. [Google Scholar] [CrossRef]
- Iqbal, U.; Hameed, M.; Ahmad, F.; Ahmad, M.S.A.; Ashraf, M.; Kaleem, M.; Irshad, M. Contribution of structural and functional modifications to wide distribution of Bermuda grass Cynodon dactylon (L.) Pers. Flora 2022, 286, 151973. [Google Scholar] [CrossRef]
- Richards, L.A. USDA Laboratory Staff, United State Department of Agriculture, Agriculture Handbook No. 60; U. S. Government Printing Office: Washington, DC, USA, 1954.
- Kowalenco, C.G.; Lowe, L.E. Determination of nitrate in soil extracts. Soil Sci. Soc. Am. Proc. 1973, 37, 660. [Google Scholar] [CrossRef]
- Yoshida, S.; Forno, D.A.; Cock, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice; IRRI: Los Banos, CA, USA, 1976. [Google Scholar]
- Schrader, J.; Shi, P.; Royer, D.L.; Peppe, D.J.; Gallagher, R.V.; Li, Y.; Wright, I.J. Leaf size estimation based on leaf length, width and shape. Ann. Bot. 2021, 128, 395–406. [Google Scholar] [CrossRef]
- Wolf, B. An improved universal extracting solution and its use for diagnosing soil fertility. Commun. Soil Sci. Plant Anal. 1982, 13, 1005–1033. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Scott, K.J. Detection and Measurement of Carotenoids by UV/VIS Spectrophotometry. Curr. Protoc. Food Anal. Chem. 2001, 1, F2.2.1–F2.2.10. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grattan, S.R.; Grieve, C.M. Betaine status in wheat in relation to nitrogen stress and to transient salinity stress. Plant Soil 1985, 85, 3–9. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive for the quantitation of microgram quantitites of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Calorimetric Dubois method for determination of sugar and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Hamilton, P.B.; Van Slyke, D.D. Amino acid determination with ninhydrin. J. Biol. Chem. 1943, 150, 231–233. [Google Scholar] [CrossRef]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: New York, NY, USA, 1999; Volume 198, p. 322. [Google Scholar]
- Naz, N.; Fatima, S.; Hameed, M.; Ashraf, M.; Ahmad, M.S.A.; Ahmad, F.; Shah, S.M.R.; Islam, F.; Ahmad, I.; Ejaz, F.; et al. Contribution of structural and functional adaptations of hyper-accumulator Suaeda vera Forssk. ex J.F. Gmel. to adaptability across salinity gradients in hot desert. Environ. Sci. Pollut. Res. 2022, 29, 64077–64095. [Google Scholar] [CrossRef]
- Steel, R.G.; Torrie, J.H.; Dickey, D. Principles and Procedures of Statistics, a Biometrical Approach; The McGraw-Hill Co., Inc.: New York, NY, USA, 1997. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cenchrus pennisetiformis | Cenchrus prieurii | Cenchrus setiger | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Normal-Control | Aridity | Salinity | Cold | Normal-Control | Aridity | Salinity | Cold | Normal-Control | Aridity | Salinity | Cold | |
| El | 91.3 | 145.8 | 201.1 | 2182.7 | 147.3 | 151.4 | 292.5 | 2316.4 | 192.3 | 108.4 | 192.5 | 2446.3 |
| ARF | 324.3 | 122.4 | 309.4 | 1590.8 | 271.5 | 145.6 | 350.3 | 1603.7 | 368.5 | 117.6 | 320.7 | 1747.2 |
| ASF | -- | -- | -- | 320 | -- | -- | -- | 335 | -- | -- | -- | 391 |
| MxT | 45.2 | 49.9 | 45.0 | 23.7 | 45.2 | 48 | 45.2 | 20.7 | 44.2 | 50.1 | 45 | 21.1 |
| MnT | 2.1 | 1.1 | 1.3 | −3.1 | 1.1 | 1.2 | 1.1 | −4.2 | 1.1 | 1.1 | 1.4 | −5.6 |
| Com | Cdd, Sba, Aja | Ajq, Lsc, Cpo | Fdi, Sva, Ala | Maf, Hna, Pro | Tdo, Ssp, Pka | Cpo, Cci, Aad | Cdd, Rst, Aad | Cde, Pwa, Tan | Cda, Cpo, Sba | Cdd, Hsa, Sim | Ala, Cla, Pka | Lpe, Acy, Pro |
| OOM | 0.82 | 0.71 | 0.71 | 1.2 | 1.1 | 0.46 | 0.88 | 1.1 | 0.91 | 0.48 | 0.55 | 0.92 |
| OSP | 25.6 | 18.4 | 24.7 | 25.2 | 28.6 | 20.8 | 25.2 | 34.7 | 29.3 | 19.0 | 32.6 | 25.1 |
| OEC | 2.1 | 2.9 | 8.5 | 1.3 | 2.3 | 2.9 | 7.7 | 1.4 | 2.8 | 15.7 | 20.4 | 1.0 |
| OpH | 7.8 | 8.1 | 7.6 | 8.1 | 8.1 | 8.0 | 8.1 | 7.6 | 7.5 | 8.6 | 8.1 | 8.1 |
| ONa | 192.3 | 296.4 | 983.5 | 147.2 | 296.3 | 278.3 | 791.7 | 136.4 | 232.6 | 2117.1 | 2512.5 | 146.7 |
| OK | 83.7 | 112.1 | 67.4 | 76.0 | 87.4 | 52.3 | 141.3 | 101.5 | 127.5 | 147.4 | 75.1 | 77.5 |
| OCa | 57.5 | 78.9 | 46.6 | 53.6 | 67.4 | 95.1 | 82.2 | 77.5 | 67.2 | 57.6 | 78.2 | 49.9 |
| OPO | 6.8 | 10.3 | 4.9 | 7.1 | 4.2 | 5.6 | 7.2 | 7.3 | 7.5 | 9.1 | 3.8 | 7.2.6 |
| ONO | 3.5 | 4.3 | 4.8 | 2.7 | 4.4 | 4.1 | 1.8 | 3.5 | 2.3 | 2.8 | 2.9 | 2.9 |
| Variables | Oel | OOM | OSP | OEC | OpH | Ona | OK | Oca | ONO | OPO |
|---|---|---|---|---|---|---|---|---|---|---|
| MPH | −0.268 | −0.176 | −0.227 | 0.160 | 0.083 | 0.258 | 0.334 | −0.361 | −0.148 | 0.053 |
| MRL | −0.039 | 0.613 | −0.334 | −0.159 | 0.074 | −0.280 | 0.100 | 0.429 | −0.546 | 0.476 |
| MSF | −0.310 | 0.709 | 0.047 | −0.091 | 0.220 | −0.107 | 0.378 | −0.043 | −0.426 | 0.371 |
| MSD | −0.303 | 0.709 | 0.044 | −0.057 | 0.260 | −0.108 | 0.396 | −0.095 | −0.447 | 0.425 |
| MRF | 0.403 | −0.351 | 0.357 | −0.295 | −0.267 | −0.271 | −0.098 | 0.339 | 0.033 | 0.274 |
| MRD | 0.425 | −0.372 | 0.391 | −0.257 | −0.298 | −0.173 | −0.125 | 0.321 | −0.063 | 0.290 |
| MLA | −0.206 | 0.113 | −0.285 | 0.230 | 0.349 | 0.274 | 0.339 | −0.188 | −0.562 | 0.353 |
| MLN | −0.367 | 0.076 | −0.086 | 0.425 | −0.061 | 0.601 | 0.259 | −0.107 | −0.706 | 0.460 |
| MIL | 0.702 | 0.300 | 0.067 | −0.150 | 0.239 | −0.054 | −0.136 | −0.256 | −0.233 | 0.078 |
| RR | −0.291 | 0.023 | −0.199 | −0.402 | −0.022 | −0.364 | 0.410 | 0.563 | 0.007 | 0.043 |
| Rep | −0.385 | 0.180 | −0.173 | 0.021 | 0.142 | −0.107 | 0.354 | 0.398 | −0.040 | 0.406 |
| Rco | 0.132 | 0.719 | −0.391 | −0.139 | −0.013 | −0.159 | −0.011 | 0.113 | −0.295 | 0.309 |
| Ren | −0.186 | 0.544 | −0.410 | −0.207 | 0.397 | −0.542 | −0.028 | 0.570 | −0.058 | 0.089 |
| RPr | 0.210 | 0.286 | −0.346 | −0.232 | −0.207 | −0.305 | 0.111 | −0.322 | 0.090 | 0.121 |
| RMx | −0.218 | 0.439 | −0.223 | −0.130 | 0.264 | −0.341 | 0.163 | 0.515 | −0.444 | 0.425 |
| RPh | −0.165 | −0.101 | −0.418 | −0.113 | −0.016 | −0.090 | 0.079 | 0.854 | −0.026 | 0.023 |
| Rae | −0.287 | 0.199 | −0.099 | −0.132 | −0.212 | −0.003 | 0.316 | −0.045 | −0.179 | 0.231 |
| RPt | −0.004 | 0.122 | −0.221 | −0.407 | −0.067 | −0.695 | 0.136 | 0.437 | 0.096 | 0.253 |
| SR | 0.125 | 0.507 | −0.256 | −0.521 | 0.007 | −0.528 | 0.063 | 0.360 | 0.027 | 0.012 |
| Sep | 0.179 | 0.334 | −0.012 | −0.092 | −0.109 | −0.370 | −0.151 | −0.559 | 0.006 | 0.088 |
| Sco | 0.216 | 0.217 | −0.052 | −0.453 | −0.274 | −0.475 | 0.090 | −0.196 | 0.043 | 0.144 |
| CMx | 0.079 | 0.309 | −0.462 | 0.738 | 0.718 | 0.373 | −0.237 | −0.227 | −0.426 | 0.539 |
| CVB | 0.202 | 0.095 | 0.206 | −0.719 | −0.531 | −0.566 | 0.203 | 0.373 | −0.011 | −0.013 |
| SSc | 0.327 | 0.453 | −0.278 | −0.534 | −0.033 | −0.545 | 0.033 | 0.163 | −0.269 | 0.091 |
| SPh | −0.025 | 0.212 | −0.108 | −0.446 | 0.130 | −0.588 | 0.098 | 0.631 | 0.269 | −0.177 |
| HT | 0.188 | 0.199 | −0.100 | 0.265 | −0.130 | −0.157 | −0.538 | 0.132 | −0.010 | 0.520 |
| Hep | 0.302 | 0.581 | −0.036 | −0.204 | −0.286 | −0.333 | −0.093 | −0.032 | −0.470 | 0.605 |
| HVB | 0.635 | 0.028 | 0.102 | −0.280 | −0.235 | −0.474 | −0.523 | 0.318 | 0.066 | 0.257 |
| HSc | 0.539 | 0.296 | −0.114 | −0.211 | −0.344 | −0.434 | −0.376 | −0.166 | 0.206 | 0.173 |
| Hco | −0.262 | 0.478 | −0.252 | 0.041 | −0.240 | −0.126 | 0.157 | 0.128 | −0.130 | 0.490 |
| Lep | −0.042 | 0.159 | −0.048 | −0.258 | −0.231 | −0.475 | −0.194 | 0.576 | 0.356 | 0.042 |
| Lco | −0.104 | −0.386 | 0.583 | −0.237 | −0.424 | −0.071 | 0.246 | −0.280 | 0.302 | −0.316 |
| LMs | −0.200 | 0.054 | 0.146 | −0.299 | 0.133 | −0.363 | 0.249 | 0.569 | 0.393 | −0.173 |
| LMx | 0.161 | 0.202 | −0.164 | −0.432 | −0.280 | −0.571 | 0.155 | −0.125 | 0.317 | 0.056 |
| LPh | 0.166 | −0.311 | 0.195 | −0.399 | −0.265 | −0.284 | −0.137 | 0.638 | 0.068 | −0.214 |
| LVB | 0.068 | 0.161 | −0.173 | −0.154 | −0.520 | −0.394 | −0.144 | −0.019 | 0.292 | 0.139 |
| LMd | −0.227 | 0.147 | −0.017 | −0.487 | −0.432 | −0.263 | 0.709 | 0.211 | −0.070 | 0.199 |
| LLm | 0.030 | 0.126 | −0.152 | −0.550 | −0.307 | −0.605 | 0.215 | 0.260 | 0.390 | −0.179 |
| LBl | 0.350 | 0.219 | −0.313 | −0.258 | −0.070 | −0.369 | −0.114 | −0.296 | 0.281 | −0.160 |
| DSD | 0.433 | 0.075 | −0.259 | 0.136 | −0.212 | −0.078 | −0.590 | −0.472 | 0.292 | −0.149 |
| BSD | 0.234 | 0.044 | −0.301 | 0.050 | −0.390 | −0.136 | −0.525 | −0.346 | 0.398 | −0.243 |
| DSA | −0.102 | −0.005 | 0.139 | −0.374 | 0.185 | −0.252 | 0.077 | 0.746 | 0.172 | −0.333 |
| BSA | 0.024 | −0.037 | 0.161 | −0.364 | 0.123 | −0.268 | −0.040 | 0.713 | 0.305 | −0.364 |
| PPt | 0.043 | 0.417 | −0.539 | 0.326 | 0.077 | −0.045 | −0.639 | 0.119 | 0.119 | −0.027 |
| PAA | −0.423 | −0.047 | −0.436 | 0.332 | −0.053 | 0.003 | −0.068 | −0.002 | 0.570 | −0.230 |
| PSg | −0.283 | −0.245 | 0.050 | 0.530 | 0.082 | 0.403 | −0.006 | 0.129 | −0.247 | 0.347 |
| PGB | 0.169 | 0.700 | −0.186 | −0.193 | −0.080 | −0.304 | −0.203 | −0.154 | 0.061 | 0.057 |
| PPl | −0.203 | 0.036 | −0.362 | 0.308 | 0.152 | 0.126 | −0.423 | 0.536 | 0.247 | −0.256 |
| Pca | −0.088 | −0.094 | 0.061 | −0.044 | −0.060 | −0.171 | −0.083 | 0.778 | 0.000 | 0.210 |
| PCb | −0.312 | −0.005 | −0.062 | 0.002 | 0.200 | 0.061 | 0.046 | 0.770 | 0.091 | −0.189 |
| PTC | −0.222 | 0.120 | 0.097 | −0.099 | 0.139 | −0.114 | −0.101 | 0.518 | 0.317 | −0.274 |
| PSN | −0.590 | 0.040 | −0.583 | 0.731 | 0.069 | 0.425 | −0.188 | −0.158 | 0.233 | −0.054 |
| PSK | −0.533 | −0.075 | −0.266 | 0.017 | 0.132 | −0.160 | 0.403 | −0.109 | 0.091 | −0.056 |
| PSC | −0.093 | 0.032 | −0.673 | 0.650 | 0.445 | 0.350 | −0.372 | 0.165 | 0.062 | 0.001 |
| PCr | 0.364 | −0.400 | 0.371 | 0.168 | −0.200 | −0.158 | −0.543 | −0.329 | 0.174 | 0.203 |
| Traits | Normal control | Drought | Salinity | Cold |
|---|---|---|---|---|
| Cenchrus pennisetiformis | ||||
| Morphology | MPH, MLA, MRF | MRL, MLN, MSF, MSD, MPH | MRL, MLA, MSF, MSD, MRF, MRD | MIL |
| Physiology | PPt, PAA, PPl, PSN | PSg, Pca, PTC | PSg, Pca, PTC, PSN | |
| Stem anatomy | SCo | SMx | SSc | |
| Leaf anatomy | LVB | Lep, LBl | Lpa, LPh | |
| Leaf sheath anatomy | HSc | HT, HPa | ||
| Root anatomy | RR, RPt | Rep, RCo, RMx | RCo | RPh |
| Cenchrus prieurii | ||||
| Morphology | MSF, MSD | MRL, MLN, MLA, MRF, MRD | MRF, MRD | |
| Physiology | PGB, PTC | PAA, PCr | PSN | PCr, PPt, PAA |
| Stem anatomy | SPh | SSc | SCo | |
| Leaf anatomy | LBl. LMd, LLm, LMx, LVB, LPA | LPA, LPh | ||
| Leaf sheath anatomy | Hpa | HSc, HT, Hpa | HT, HVB | |
| Root anatomy | Rae, RPt, RPh | RPr, RPt, RR | RMx, Rep | RCo |
| Cenchrus setiger | ||||
| Morphology | MLA | MRL | ||
| Physiology | PPt | PSN, PCr | PCr | |
| Stem anatomy | SPh | SMx, Sep, SVB, SPh | SPh | Sep, SSc |
| Leaf anatomy | LPA, LMd, LMS, LMx, LVB, LBl | LBl, LMd, LPA, LPh, LVB | ||
| Leaf sheath anatomy | HT, Hpa, HVB | HT | ||
| Root anatomy | Rco | RPh | Rae, RPt | Ren, Rco, RPt Rsp, RMx |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basharat, S.; Ahmad, F.; Hameed, M.; Ahmad, M.S.A.; Asghar, A.; Fatima, S.; Ahmad, K.S.; Shah, S.M.R.; Hashem, A.; Avila-Quezada, G.D.; et al. Structural and Functional Strategies in Cenchrus Species to Combat Environmental Extremities Imposed by Multiple Abiotic Stresses. Plants 2024, 13, 203. https://doi.org/10.3390/plants13020203
Basharat S, Ahmad F, Hameed M, Ahmad MSA, Asghar A, Fatima S, Ahmad KS, Shah SMR, Hashem A, Avila-Quezada GD, et al. Structural and Functional Strategies in Cenchrus Species to Combat Environmental Extremities Imposed by Multiple Abiotic Stresses. Plants. 2024; 13(2):203. https://doi.org/10.3390/plants13020203
Chicago/Turabian StyleBasharat, Sana, Farooq Ahmad, Mansoor Hameed, Muhammad Sajid Aqeel Ahmad, Ansa Asghar, Sana Fatima, Khawaja Shafique Ahmad, Syed Mohsan Raza Shah, Abeer Hashem, Graciela Dolores Avila-Quezada, and et al. 2024. "Structural and Functional Strategies in Cenchrus Species to Combat Environmental Extremities Imposed by Multiple Abiotic Stresses" Plants 13, no. 2: 203. https://doi.org/10.3390/plants13020203