
Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea
 , ,
, ,  ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Variation in Soil Properties by Land-Use Type
2.2. Edaphic Drivers of Fine-Root Mass
2.3. Variations in Standing Fine-Root Biomass and Necromass across Four Land-Use Types
2.4. Vertical Distribution of Fine-Root Biomass and Necromass in Four Land-Use Types
3. Discussion
3.1. Variation in Soil Physical and Chemical Properties across Land-Use Types
3.2. Variation in Standing Root Mass and Fine-Root Vertical Distribution across Land-Use Types
3.3. Implications for Sustainable Urban Planning
4. Materials and Methods
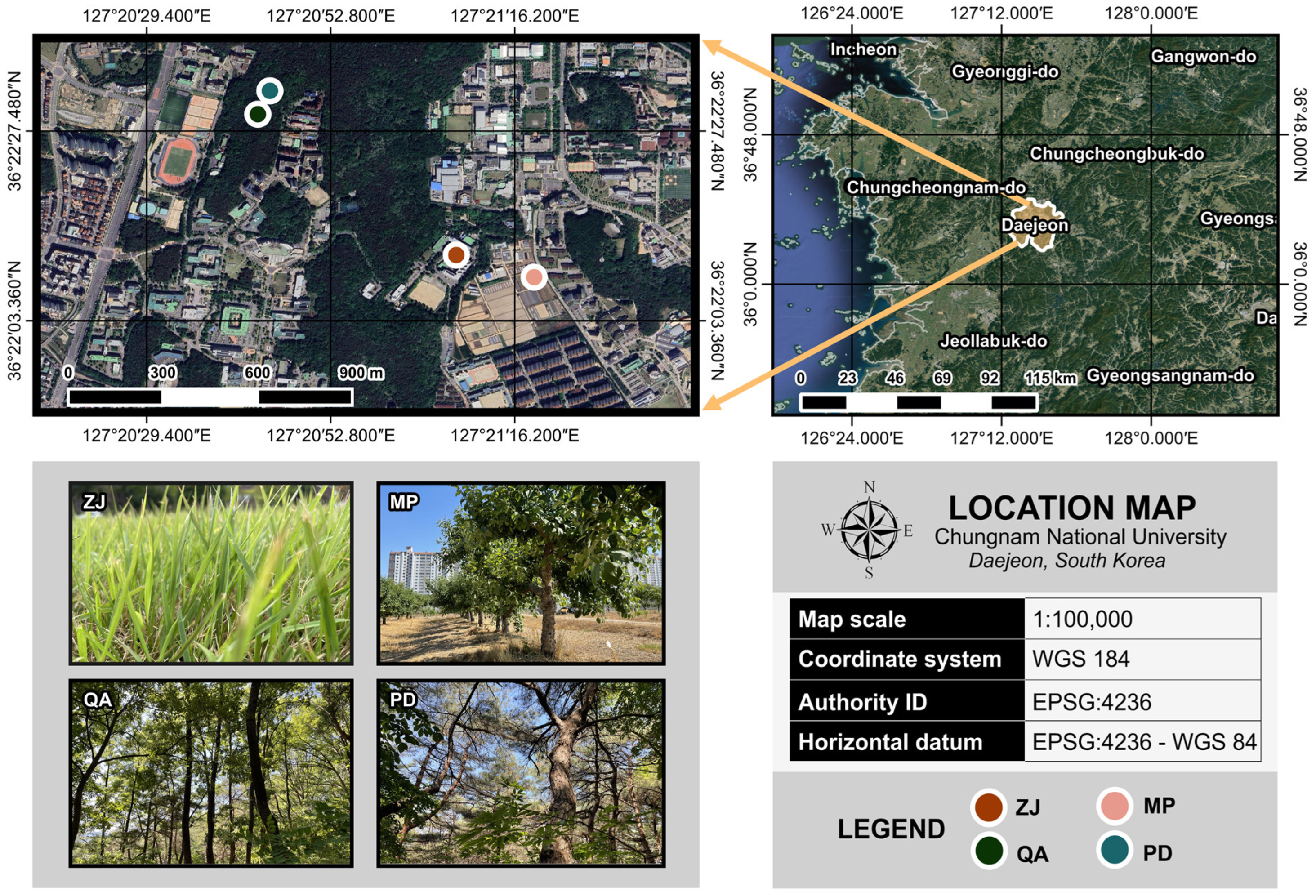
4.1. Study Site
4.2. Soil Sampling
4.3. Fine-Root Mass Inventory
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Creutzig, F.; d’Amour, C.B.; Weddige, U.; Fuss, S.; Beringer, T.; Gläser, A.; Kalkuhl, M.; Steckel, J.C.; Radebach, A.; Edenhofer, O. Assessing Human and Environmental Pressures of Global Land-Use Change 2000–2010. Glob. Sustain. 2019, 2, e1. [Google Scholar] [CrossRef]
- United Nations. World Urbanization Prospects, the 2011 Revision. In Population Division, Department of Economic and Social Affairs, United Nations Secretariat; United Nations: New York, NY, USA, 2014. [Google Scholar]
- Wei, H.; Guo, P.; Zheng, H.; He, X.; Wang, P.; Ren, Z.; Zhai, C. Micro-Scale Heterogeneity in Urban Forest Soils Affects Fine Root Foraging by Ornamental Seedlings of Buddhist Pine and Northeast Yew. Urban For. Urban Green. 2017, 28, 63–72. [Google Scholar] [CrossRef]
- Upadhyay, S.; Raghubanshi, A.S. Chapter 16—Determinants of Soil Carbon Dynamics in Urban Ecosystems; Verma, P., Singh, P., Singh, R., Raghubanshi, A.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 299–314. ISBN 978-0-12-820730-7. [Google Scholar]
- IPCC. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- DeFries, R.S.; Foley, J.A.; Asner, G.P. Land-Use Choices: Balancing Human Needs and Ecosystem Function. Front. Ecol. Environ. 2004, 2, 249–257. [Google Scholar] [CrossRef]
- Leul, Y.; Assen, M.; Damene, S.; Legass, A. Effects of Land Use Types on Soil Quality Dynamics in a Tropical Sub-Humid Ecosystem, Western Ethiopia. Ecol. Indic. 2023, 147, 110024. [Google Scholar] [CrossRef]
- Ramesh, T.; Bolan, N.S.; Kirkham, M.B.; Wijesekara, H.; Kanchikerimath, M.; Rao, C.S.; Sandeep, S.; Rinklebe, J.; Ok, Y.S.; Choudhury, B.U. Soil Organic Carbon Dynamics: Impact of Land Use Changes and Management Practices: A Review. Adv. Agron. 2019, 156, 1–107. [Google Scholar]
- Cornejo, N.S.; Becker, J.N.; Hemp, A.; Hertel, D. Effects of Land-Use Change and Disturbance on the Fine Root Biomass, Dynamics, Morphology, and Related C and N Fluxes to the Soil of Forest Ecosystems at Different Elevations at Mt. Kilimanjaro (Tanzania). Oecologia 2023, 201, 1089–1107. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-G.; Kirschbaum, M.U.F.; Eichler-Löbermann, B.; Gifford, R.M.; Liáng, L.L. The Effect of Land-Use Change on Soil C, N, P, and Their Stoichiometries: A Global Synthesis. Agric. Ecosyst. Environ. 2023, 348, 108402. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.-D. A Global Budget for Fine Root Biomass, Surface Area, and Nutrient Contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef]
- Chapin, F.S.; Eviner, V.T. 8.06—Biogeochemistry of Terrestrial Net Primary Production. In Treatise on Geochemistry, 2nd ed.; Holland, H.D., Turekian, K.K., Eds.; Elsevier: Oxford, UK, 2007; pp. 1–35; 189–216. ISBN 978-0-08-043751-4. [Google Scholar]
- Van Do, T.; Sato, T.; Saito, S.; Kozan, O. Fine-Root Production and Litterfall: Main Contributions to Net Primary Production in an Old-Growth Evergreen Broad-Leaved Forest in Southwestern Japan. Ecol. Res. 2015, 30, 921–930. [Google Scholar] [CrossRef]
- Norby, R.J.; Jackson, R.B. Root Dynamics and Global Change: Seeking an Ecosystem Perspective. New Phytol. 2000, 147, 3–12. [Google Scholar] [CrossRef]
- Sims, P.L.; Singh, J.S. The Structure and Function of Ten Western North American Grasslands: IV. Compartmental Transfers and Energy Flow Within the Ecosystem. J. Ecol. 1978, 66, 983–1009. [Google Scholar] [CrossRef]
- Dong, L.; Berg, B.; Sun, T.; Wang, Z.; Han, X. Response of Fine Root Decomposition to Different Forms of N Deposition in a Temperate Grassland. Soil Biol. Biochem. 2020, 147, 107845. [Google Scholar] [CrossRef]
- Zanotelli, D.; Montagnani, L.; Manca, G.; Scandellari, F.; Tagliavini, M. Net Ecosystem Carbon Balance of an Apple Orchard. Eur. J. Agron. 2015, 63, 97–104. [Google Scholar] [CrossRef]
- Gan, Z.; Zhou, Z.; Liu, W. Vertical Distribution and Seasonal Dynamics of Fine Root Parameters for Apple Trees of Different Ages on the Loess Plateau of China. Agric. Sci. China 2010, 9, 46–55. [Google Scholar] [CrossRef]
- Coll, L.; Camarero, J.J.; Martinez De Aragon, J. Fine Root Seasonal Dynamics, Plasticity, and Mycorrhization in 2 Coexisting Mediterranean Oaks with Contrasting Aboveground Phenology. Ecoscience 2012, 19, 238–245. [Google Scholar] [CrossRef]
- Hao, H.; Wei, Y.; Cao, D.; Guo, Z.; Shi, Z. Vegetation Restoration and Fine Roots Promote Soil Infiltrability in Heavy-Textured Soils. Soil Tillage Res. 2020, 198, 104542. [Google Scholar] [CrossRef]
- Huang, G.; Zhao, X.; Su, Y.; Zhao, H.; Zhang, T. Vertical Distribution, Biomass, Production and Turnover of Fine Roots along a Topographical Gradient in a Sandy Shrubland. Plant Soil 2008, 308, 201–212. [Google Scholar] [CrossRef]
- Xie, C.; Cai, S.; Yu, B.; Yan, L.; Liang, A.; Che, S. The Effects of Tree Root Density on Water Infiltration in Urban Soil Based on a Ground Penetrating Radar in Shanghai, China. Urban For. Urban Green. 2020, 50, 126648. [Google Scholar] [CrossRef]
- Kotowska, M.M.; Leuschner, C.; Triadiati, T.; Hertel, D. Conversion of Tropical Lowland Forest Reduces Nutrient Return through Litterfall, and Alters Nutrient Use Efficiency and Seasonality of Net Primary Production. Oecologia 2016, 180, 601–618. [Google Scholar] [CrossRef]
- Pransiska, Y.; Triadiati, T.; Tjitrosoedirjo, S.; Hertel, D.; Kotowska, M.M. Forest Conversion Impacts on the Fine and Coarse Root System, and Soil Organic Matter in Tropical Lowlands of Sumatera (Indonesia). For. Ecol. Manag. 2016, 379, 288–298. [Google Scholar] [CrossRef]
- Ishaq, M.; Ibrahim, M.; Lal, R. Tillage Effects on Soil Properties at Different Levels of Fertilizer Application in Punjab, Pakistan. Soil Tillage Res. 2002, 68, 93–99. [Google Scholar] [CrossRef]
- Ryadin, A.R.; Janz, D.; Schneider, D.; Tjoa, A.; Irawan, B.; Daniel, R.; Polle, A. Early Effects of Fertilizer and Herbicide Reduction on Root-Associated Biota in Oil Palm Plantations. Agronomy 2022, 12, 199. [Google Scholar] [CrossRef]
- Phillips, R.P.; Fahey, T.J. Fertilization Effects on Fineroot Biomass, Rhizosphere Microbes and Respiratory Fluxes in Hardwood Forest Soils. New Phytol. 2007, 176, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, K.; Shibata, H.; Takagi, K.; Satoh, F.; Koike, T.; Sasa, K. Temporal Variation in Fine-Root Biomass, Production and Mortality in a Cool Temperate Forest Covered with Dense Understory Vegetation in Northern Japan. For. Ecol. Manag. 2013, 310, 700–710. [Google Scholar] [CrossRef]
- Solly, E.; Schöning, I.; Boch, S.; Müller, J.; Socher, S.A.; Trumbore, S.E.; Schrumpf, M. Mean Age of Carbon in Fine Roots from Temperate Forests and Grasslands with Different Management. Biogeosciences 2013, 10, 4833–4843. [Google Scholar] [CrossRef]
- Raizada, A.; Jayaprakash, J.; Rathore, A.C.; Tomar, J.M.S. Distribution of Fine Root Biomass of Fruit and Forest Tree Species Raised on Old River Bed Lands in the North West Himalaya. Trop. Ecol. 2013, 54, 251–261. [Google Scholar]
- Leuschner, C.; Harteveld, M.; Hertel, D. Consequences of Increasing Forest Use Intensity for Biomass, Morphology and Growth of Fine Roots in a Tropical Moist Forest on Sulawesi, Indonesia. Agric. Ecosyst. Environ. 2009, 129, 474–481. [Google Scholar] [CrossRef]
- Magalhães, T.M.; Mamugy, F.P.S. Fine Root Biomass and Soil Properties Following the Conversion of Miombo Woodlands to Shifting Cultivation Lands. Catena 2020, 194, 104693. [Google Scholar] [CrossRef]
- Pregitzer, K.S. Fine Roots of Trees: A New Perspective. New Phytol. 2002, 154, 267–270. [Google Scholar] [CrossRef]
- Hertel, D.; Harteveld, M.A.; Leuschner, C. Conversion of a Tropical Forest into Agroforest Alters the Fine Root-Related Carbon Flux to the Soil. Soil Biol. Biochem. 2009, 41, 481–490. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Lehto, T.; Makkonen, K. Fine Roots and Soil Properties BT—Forest Condition in a Changing Environment: The Finnish Case; Mälkönen, E., Ed.; Springer: Dordrecht, The Netherlands, 2000; pp. 203–217. ISBN 978-94-015-9373-1. [Google Scholar]
- Bengough, A.G.; Mullins, C.E. Mechanical Impedance to Root Growth: A Review of Experimental Techniques and Root Growth Responses. J. Soil Sci. 1990, 41, 341–358. [Google Scholar] [CrossRef]
- Silver, W.L.; Neff, J.; McGroddy, M.; Veldkamp, E.; Keller, M.; Cosme, R. Effects of Soil Texture on Belowground Carbon and Nutrient Storage in a Lowland Amazonian Forest Ecosystem. Ecosystems 2000, 3, 193–209. [Google Scholar] [CrossRef]
- Barrow, N.J.; Hartemink, A.E. The Effects of PH on Nutrient Availability Depend on Both Soils and Plants. Plant Soil 2023, 487, 21–37. [Google Scholar] [CrossRef]
- Chen, L.; Mu, X.; Yuan, Z.; Deng, Q.; Chen, Y.; Yuan, L.Y.; Ryan, L.T.; Kallenbach, R.L. Soil Nutrients and Water Affect the Age-Related Fine Root Biomass but Not Production in Two Plantation Forests on the Loess Plateau, China. J. Arid Environ. 2016, 135, 173–180. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Fine Root Biomass, Production, Turnover Rates, and Nutrient Contents in Boreal Forest Ecosystems in Relation to Species, Climate, Fertility, and Stand Age: Literature Review and Meta-Analyses. CRC. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Neina, D. The Role of Soil PH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Sun, Y.; Bi, H.; Xu, H.; Duan, H.; Peng, R.; Wang, J. Variation of Fine Roots Distribution in Apple (Malus Pumila M.)–Crop Intercropping Systems on the Loess Plateau of China. Agronomy 2018, 8, 280. [Google Scholar] [CrossRef]
- Ge, S.; Zhu, Z.; Jiang, Y. Long-Term Impact of Fertilization on Soil PH and Fertility in an Apple Production System. J. Soil Sci. Plant Nutr. 2018, 18, 282–293. [Google Scholar] [CrossRef]
- Han, S.H.; Kim, S.; Chang, H.; Kim, H.-J.; Khamzina, A.; Son, Y. Soil Depth- and Root Diameter-Related Variations Affect Root Decomposition in Temperate Pine and Oak Forests. J. Plant Ecol. 2019, 12, 871–881. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; Agronomy Monographs; Wiley: Hoboken, NJ, USA, 1983; pp. 539–579. ISBN 9780891189770. [Google Scholar]
- Park, B.B.; Rahman, A.; Han, S.H.; Youn, W.B.; Hyun, H.J.; Hernandez, J.; An, J.Y. Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea. Forests 2020, 11, 143. [Google Scholar] [CrossRef]
- Uselman, S.M.; Qualls, R.G.; Lilienfein, J. Fine Root Production across a Primary Successional Ecosystem Chronosequence at Mt. Shasta, California. Ecosystems 2007, 10, 703–717. [Google Scholar] [CrossRef]
- Zhou, G.; Meng, S.; Yu, J.; Zhou, H.; Liu, Q. Quantitative Relationships between Fine Roots and Stand Characteristics. Eur. J. For. Res. 2018, 137, 385–399. [Google Scholar] [CrossRef]
- Osono, T.; Azuma, J.; Hirose, D. Plant Species Effect on the Decomposition and Chemical Changes of Leaf Litter in Grassland and Pine and Oak Forest Soils. Plant Soil 2014, 376, 411–421. [Google Scholar] [CrossRef]
- Vang-Petersen, O. Calcium Nutrition of Apple Trees: A Review. Sci. Hortic. 1980, 12, 1–9. [Google Scholar] [CrossRef]
- Park, G.-S.; Kang, G.-N.; Lee, S.-J.; Lee, H.-G.; Lee, S.; Chae, H.-M.; Ohga, S. Net Fine Root Carbon Production in Pinus Densiflora, Pinus Koraiensis, Larix Leptolepis and Quercus Acutissima Stands, Gongju Area, Chungnam Province, Korea; Faculty of Agriculture, Kyushu University: Fukuoka, Japan, 2010. [Google Scholar]
- Han, S.H.; Kim, S.; Chang, H.; Kim, H.; An, J.; Son, Y. Fine Root Biomass and Production Regarding Root Diameter in Pinus Densiflora and Quercus Serrata Forests: Soil Depth Effects and the Relationship with Net Primary Production. Turk. J. Agric. For. 2021, 45, 46–54. [Google Scholar]
- Bueis, T.; Bravo, F.; Pando, V.; Kissi, Y.-A.; Turrión, M.-B. Phosphorus Availability in Relation to Soil Properties and Forest Productivity in Pinus sylvestris L. Plantations. Ann. For. Sci. 2019, 76, 97. [Google Scholar] [CrossRef]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. An Introduction to Nutrient Management. In Soil Fertility and Fertilizers, 6th ed.; Prentice Hall Up: Saddle River, NJ, USA, 1999; p. 499. [Google Scholar]
- Joslin, J.D.; Gaudinski, J.B.; Torn, M.S.; Riley, W.J.; Hanson, P.J. Fine-Root Turnover Patterns and Their Relationship to Root Diameter and Soil Depth in a 14C-Labeled Hardwood Forest. New Phytol. 2006, 172, 523–535. [Google Scholar] [CrossRef]
- Du, H.; Liu, L.; Su, L.; Zeng, F.; Wang, K.; Peng, W.; Zhang, H.; Song, T. Seasonal Changes and Vertical Distribution of Fine Root Biomass During Vegetation Restoration in a Karst Area, Southwest China. Front. Plant Sci. 2019, 9, 2001. [Google Scholar] [CrossRef]
- Wang, C.; Han, S.; Zhou, Y.; Yan, C.; Cheng, X.; Zheng, X.; Li, M.-H. Responses of Fine Roots and Soil N Availability to Short-Term Nitrogen Fertilization in a Broad-Leaved Korean Pine Mixed Forest in Northeastern China. PLoS ONE 2012, 7, e31042. [Google Scholar] [CrossRef]
- Von Haden, A.C.; Dornbush, M.E. Depth Distributions of Belowground Production, Biomass and Decomposition in Restored Tallgrass Prairie. Pedosphere 2019, 29, 457–467. [Google Scholar] [CrossRef]
- Boeri, P.A.; Unruh, J.B.; Kenworthy, K.E.; Trenholm, L.E.; Rios, E.F. Aboveground and Belowground Traits of Turf-type Bahiagrass (Paspalum notatum Flügge) Genotypes under Simulated Drought. Int. Turfgrass Soc. Res. J. 2022, 14, 276–287. [Google Scholar] [CrossRef]
- Fu, J.; Fry, J.; Huang, B. Growth and Carbon Metabolism of Tall Fescue and Zoysiagrass as Affected by Deficit Irrigation. HortScience 2007, 42, 378–381. [Google Scholar] [CrossRef]
- Song, X.; Gao, X.; Dyck, M.; Zhang, W.; Wu, P.; Yao, J.; Zhao, X. Soil Water and Root Distribution of Apple Tree (Malus Pumila Mill) Stands in Relation to Stand Age and Rainwater Collection and Infiltration System (RWCI) in a Hilly Region of the Loess Plateau, China. Catena 2018, 170, 324–334. [Google Scholar] [CrossRef]
- Li, X.; Minick, K.J.; Li, T.; Williamson, J.C.; Gavazzi, M.; McNulty, S.; King, J.S. An Improved Method for Quantifying Total Fine Root Decomposition in Plantation Forests Combining Measurements of Soil Coring and Minirhizotrons with a Mass Balance Model. Tree Physiol. 2020, 40, 1466–1473. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Xiang, W.; Lei, P.; Deng, X.; Tian, D.; Fang, X.; Peng, C. Standing Fine Root Mass and Production in Four Chinese Subtropical Forests along a Succession and Species Diversity Gradient. Plant Soil 2014, 376, 445–459. [Google Scholar] [CrossRef]
- Jiang, P.; Wang, H.; Fu, X.; Dai, X.; Kou, L.; Wang, J. Elaborate Differences between Trees and Understory Plants in the Deployment of Fine Roots. Plant Soil 2018, 431, 433–447. [Google Scholar] [CrossRef]
- Meinen, C.; Hertel, D.; Leuschner, C. Biomass and Morphology of Fine Roots in Temperate Broad-Leaved Forests Differing in Tree Species Diversity: Is There Evidence of below-Ground Overyielding? Oecologia 2009, 161, 99–111. [Google Scholar] [CrossRef]
- Bourne, K.S.; Conway, T.M. The Influence of Land Use Type and Municipal Context on Urban Tree Species Diversity. Urban Ecosyst. 2014, 17, 329–348. [Google Scholar] [CrossRef]
- Jim, C.Y. Sustainable Urban Greening Strategies for Compact Cities in Developing and Developed Economies. Urban Ecosyst. 2013, 16, 741–761. [Google Scholar] [CrossRef]
- Francos, M.; Bogunovic, I.; Úbeda, X.; Pereira, P. Soil Physico-Chemical Properties and Organic Carbon Stocks across Different Land Use in an Urban Park of Vilnius, Lithuania. J. Cent. Eur. Agric. 2023, 24, 519–530. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Physical Properties | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sites | Sand | Silt | Clay | Soil Texture | SBD | |||||||||
| ZJ | a | b | c | ab | ||||||||||
| 0–10 cm | 87 (0) | 10 (0) | 3 (0.01) | sand | 1.27 (0.07) | |||||||||
| 10–30 cm | 85 (0.02) | 11 (0.02) | 4 (0) | loamy sand | 1.41 (0.07) | |||||||||
| MP | c | a | b | a | ||||||||||
| 0–10 cm | 65 (0.01) | 27 (0.01) | 8 (0.01) | sandy loam | 1.52 (0.09) | |||||||||
| 10–30 cm | 67 (0.01) | 25 (0.01) | 8 (0) | sandy loam | 1.58 (0.07) | |||||||||
| QA | b | b | b | ab | ||||||||||
| 0–10 cm | 78 (0.01) | 13 (0.01) | 9 (0.01) | sandy loam | 1.43 (0.07) | |||||||||
| 10–30 cm | 73 (0.03) | 16 (0.01) | 11 (0.02) | sandy loam | 1.45 (0.09) | |||||||||
| PD | ab | b | a | b | ||||||||||
| 0–10 cm | 73 (0.05) | 13 (0.02) | 13 (0.02) | sandy loam | 1.23 (0.06) | |||||||||
| 10–30 cm | 69 (0.04) | 15 (0.03) | 16 (0.01) | sandy loam | 1.37 (0.09) | |||||||||
| Chemical Properties | ||||||||||||||
| Exchangeable Cations | ||||||||||||||
| pH | EC | OM | AP | TN | C/N | NH4+-N | NO3−-N | CEC | K+ | Na+ | Ca2+ | Mg2+ | Al3+ | |
| ZJ | a | c | c | b | a | b | a | a | b | a | ||||
| 0–10 cm | 6.23 | 0.07 | 2.66 | 121.2 | 0.1 | 12.24 | 2.06 | 16.86 | 3.74 | 0.36 | 0.12 | 2.28 | 0.57 | 0.1 |
| (0.07) | (0.01) | (0.43) | (51.2) | (0.01) | (0.88) | (0.13) | (3.04) | (0.18) | (0.01) | (0.02) | (0.22) | (0.04) | (0.00) | |
| 10–30 cm | 6.23 | 0.05 | 0.91 | 39.4 | 0.02 | 14.16 | 1.73 | 10.23 | 4.41 | 0.23 | 0.09 | 3.19 | 0.56 | 0.1 |
| (0.06) | (0.00) | (0.14) | (25.3) | (0.00) | (1.49) | (0.34) | (2.25) | (0.46) | (0.07) | (0.03) | (0.09) | (0.12) | (0.00) | |
| MP | a | bc | c | b | b | b | a | a | a | a | ||||
| 0–10 cm | 6.9 | 0.23 | 0.23 | 296.5 | 0.06 | 10.63 | 2.25 | 7.6 | 4.92 | 0.31 | 0.08 | 3.31 | 1.25 | 0.1 |
| (0.48) | (0.07) | (0.07) | (240.6) | (0.02) | (0.92) | (0.20) | (2.38) | (0.99) | (0.10) | (0.01) | (0.68) | (0.25) | (0.00) | |
| 10–30 cm | 6.85 | 0.13 | 0.13 | 62.6 | 0.04 | 10.58 | 2.29 | 4.3 | 4.4 | 0.27 | 0.12 | 2.86 | 1.1 | 0.1 |
| (0.46) | (0.02) | (0.02) | (29.7) | (0.01) | (0.94) | (0.34) | (0.34) | (0.20) | (0.05) | (0.03) | (0.06) | (0.06) | (0.00) | |
| QA | b | ab | b | a | b | a | b | b | c | b | ||||
| 0–10 cm | 4.4 | 0.34 | 4.16 | 23.1 | 0.07 | 38.65 | 3.54 | 3.94 | 7.88 | 0.08 | 0.33 | 0.63 | 0.05 | 0.04 |
| (0.16) | (0.08) | (0.71) | (0.8) | (0.02) | (5.96) | (1.10) | (1.22) | (0.53) | (0.00) | (0.04) | (0.05) | (0.00) | (0.01) | |
| 10–30 cm | 4.55 | 0.28 | 4.22 | 12.4 | 0.06 | 49.22 | 3.1 | 3.45 | 7.2 | 0.1 | 0.41 | 0.62 | 0.05 | 0.03 |
| (0.07) | (0.06) | (0.89) | (1.0) | (0.03) | (10.41) | (1.34) | (1.49) | (1.13) | (0.02) | (0.17) | (0.07) | (0.00) | (0.01) | |
| PD | b | a | a | a | b | ab | b | c | c | a | ||||
| 0–10 cm | 4.18 | 0.46 | 6.86 | 27.2 | 0.11 | 39.45 | 5.29 | 5.92 | 5.82 | 0.09 | 0.21 | 0.44 | 0.05 | 0.12 |
| (0.06) | (0.04) | (0.73) | (1.0) | (0.02) | (3.72) | (0.89) | (0.99) | (1.56) | (0.01) | (0.12) | (0.09) | (0.00) | (0.02) | |
| 10–30 cm | 4.36 | 0.33 | 5.32 | 24.0 | 0.05 | 70.05 | 2.29 | 2.39 | 4.56 | 0.09 | 0.14 | 0.32 | 0.06 | 0.05 |
| (0.05) | (0.04) | (0.79) | (8.5) | (0.01) | (2.55) | (0.42) | (0.48) | (1.68) | (0.02) | (0.07) | (0.12) | (0.01) | (0.01) | |
| Site | Biomass (g m−2) | Necromass (g m−2) | |||||
|---|---|---|---|---|---|---|---|
| <2 mm | 2–5 mm | <5 mm | <2 mm | 2–5 mm | <5 mm | ||
| Fine root mass | |||||||
| ZJ | 246.24 (17.23) a | 193.85 (50.69) a | 440.09 (65.28) ab | 7.18 (1.90) bc | 81.31 (25.17) a | 88.49 (25.29) a | |
| MP | 48.25 (21.28) b | 50.05 (32.2) a | 98.30 (47.48) b | 1.06 (0.57) c | 0 b | 1.06 (0.57) b | |
| QA | 404.87 (56.82) a | 295.43 (117.81) a | 700.30 (153.69) a | 45.46 (9.51) a | 19.04 (14.22) b | 64.49 (8.59) a | |
| PD | 366.77 (55.10) a | 120.44 (30.91) a | 487.21 (57.35) a | 25.62 (7.17) ab | 8.71 (6.01) b | 34.33 (12.62) ab | |
| ANOVA Summary | |||||||
| Variation source | df | <2 mm | 2–5 mm | <5 mm | <2 mm | 2–5 mm | <5 mm |
| Site | 3 | <0.0001 | 0.054 | <0.001 | <0.0001 | 0.001 | 0.001 |
| Soil depth | 5 | <0.0001 | 0.002 | <0.001 | <0.0001 | <0.001 | <0.001 |
| Site x soil depth | 15 | <0.0001 | 0.003 | <0.001 | <0.0001 | <0.001 | <0.001 |
| Site | ZJ | MP | QA | PD |
|---|---|---|---|---|
| Tree density (tree ha−1) | NA | 700 | 900 | 800 |
| Mean height (m) | 3.7 | 13.8 | 12.5 | |
| Mean DBH (cm) | 15.5 | 30.3 | 24.2 | |
| Basal area (m2 ha−1) | 13.4 | 43.7 | 39.4 | |
| Dominant species | Zoysia japonica | Malus pumila | Quercus acutissima | Pinus densiflora |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, L.T.N.; An, J.Y.; Carayugan, M.B.; Hernandez, J.O.; Rahman, S.A.; Youn, W.B.; Carvalho, J.I.; Jo, M.S.; Han, S.H.; Nguyen, H.-H.; et al. Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea. Plants 2024, 13, 164. https://doi.org/10.3390/plants13020164
Tran LTN, An JY, Carayugan MB, Hernandez JO, Rahman SA, Youn WB, Carvalho JI, Jo MS, Han SH, Nguyen H-H, et al. Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea. Plants. 2024; 13(2):164. https://doi.org/10.3390/plants13020164
Chicago/Turabian StyleTran, Lan Thi Ngoc, Ji Young An, Mark Bryan Carayugan, Jonathan O. Hernandez, SK Abidur Rahman, Woo Bin Youn, Julia Inacio Carvalho, Min Seon Jo, Si Ho Han, Hai-Hoa Nguyen, and et al. 2024. "Fine-Root Distribution and Soil Physicochemical Property Variations in Four Contrasting Urban Land-Use Types in South Korea" Plants 13, no. 2: 164. https://doi.org/10.3390/plants13020164