
Ecogeography of Dioscorea remotiflora Kunth: An Endemic Species from Mexico
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Selection of Environmental Variables
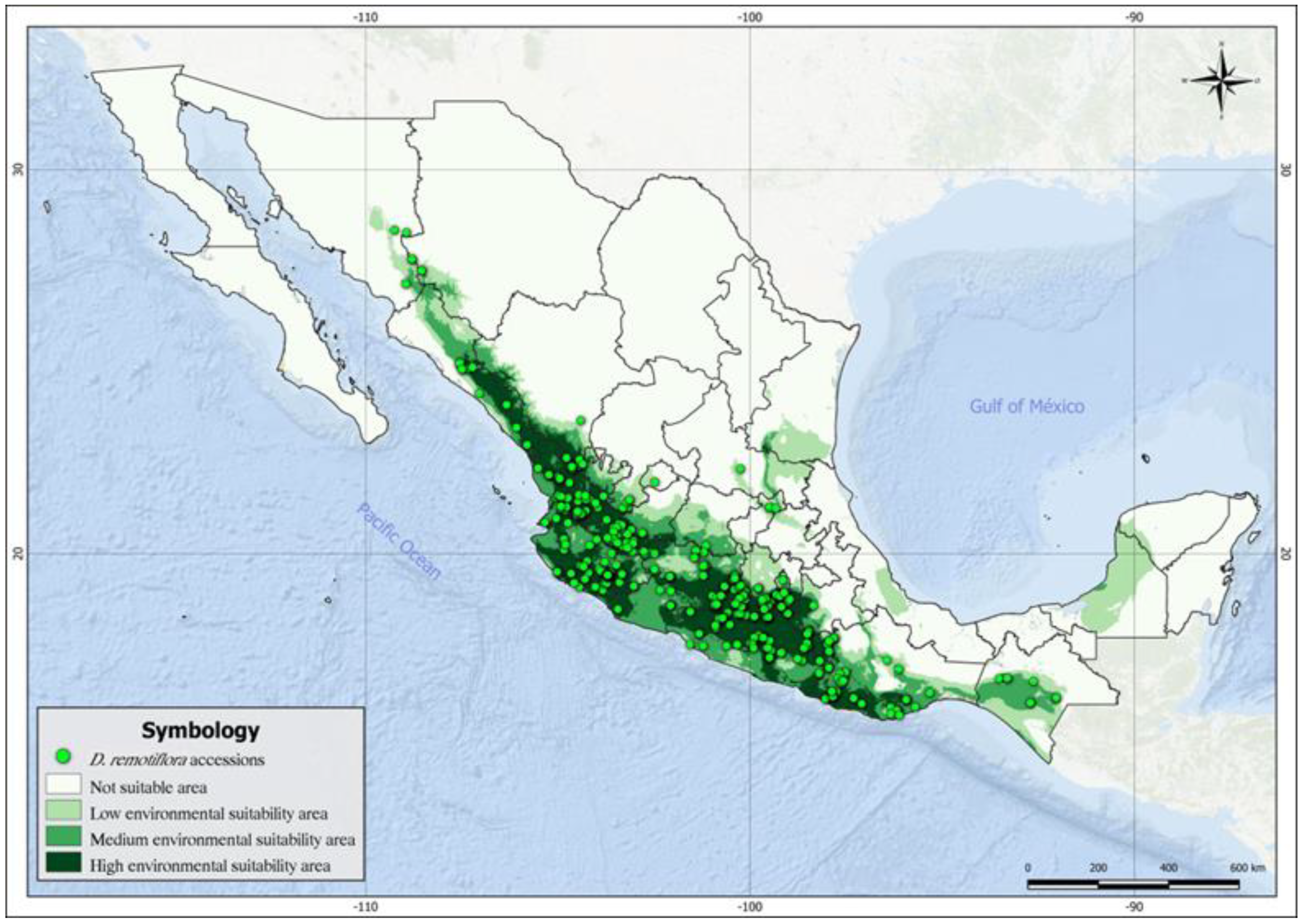
2.2. Current Distribution, Climatic Adaptation, and Ecological Descriptors
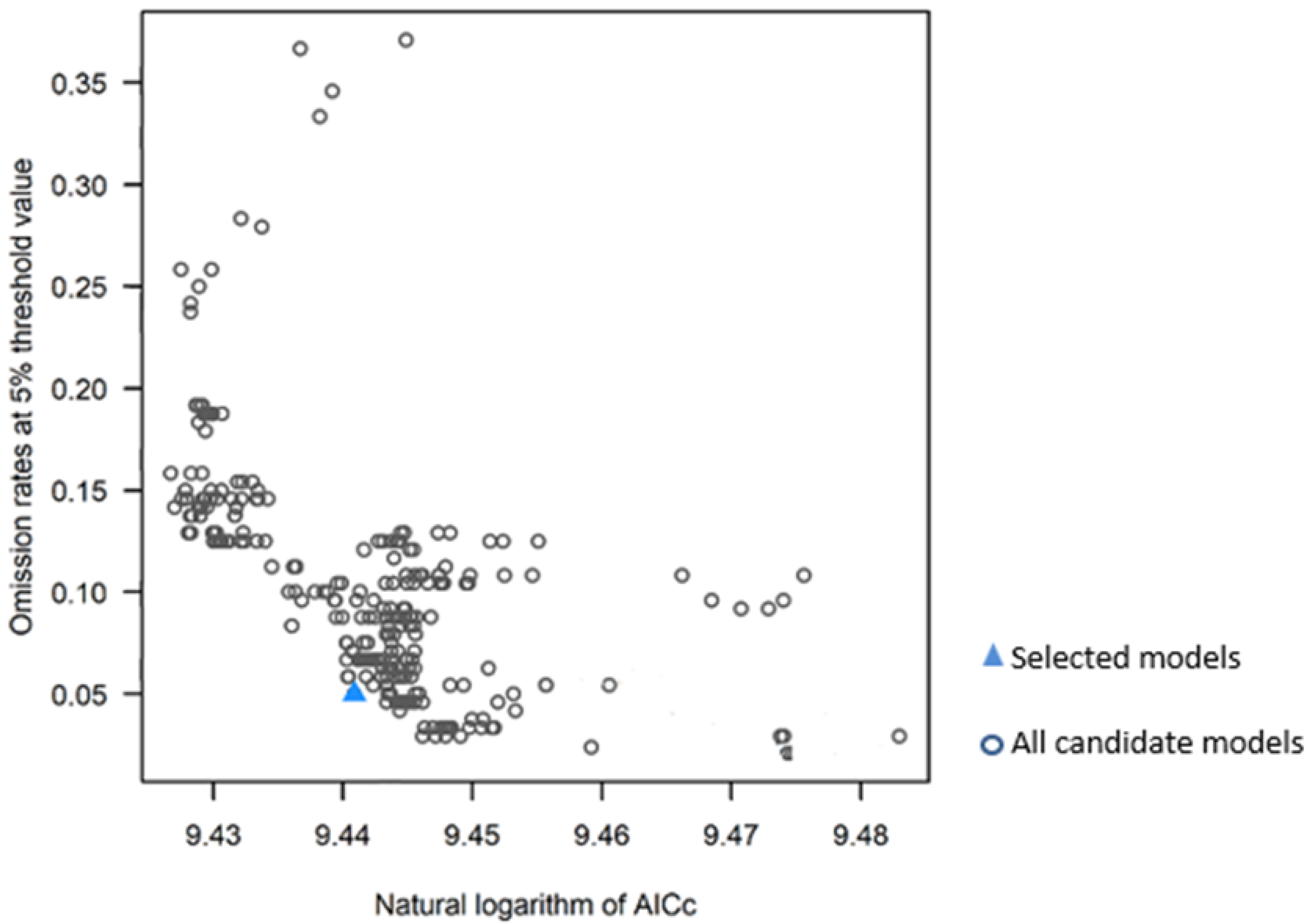
2.3. Modeling Distribution Niches of D. remotiflora
3. Discussion
3.1. Current Distribution, Climate Adaptation, and Ecological Descriptors
3.2. Modeling of Distribution Niches of D. remotiflora
3.3. Dioscorea remotiflora Cultivation Prospects
4. Materials and Methods
4.1. Occurrence Data
4.2. Climatic Data
4.3. Environmental Characterization of the Occurrence Sites
4.4. Selection of Environmental Variables
4.5. Characterization of the Adaptive Capacity of D. remotiflora
4.6. Ecological Niche Modeling
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hussein, I.; Mengs, B.; Matiwos, T. Effect of Plant Growth Regulators on in Vitro Propagation of Yam Landraces (Dioscorea Species) Using Nodal Segments. J. Biol. Agric. Healthc. 2018, 8, 13–23. Available online: https://core.ac.uk/download/pdf/234662712.pdf (accessed on 23 March 2023).
- Rodríguez, R.R.; Téllez, V.O. Las Dioscoreas (Dioscoreaceae) del Estado de Morelos, México. Anales del Instituto de Biología. Ser. Botánica 1992, 63, 67–99. Available online: https://www.redalyc.org/pdf/400/40063104.pdf (accessed on 10 March 2023).
- Castañeda, N.J.J.; Santacruz, R.F.; de Jesús, S.G.J.; Parra, J.R.; De la Cruz, L.L.; Barba, R. Shading and Container Effects on the Weight of the Dioscorea sparsiflora Tuber. Agron. J. 2017, 109, 33–38. [Google Scholar] [CrossRef]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Dioscorea remotiflora. Available online: https://enciclovida.mx/especies/192190-dioscorea-remotiflora (accessed on 9 September 2022).
- Obidiegwu, J.E.; Akpabio, E.M. The geography of yam cultivation in southern Nigeria: Exploring its social meanings and cultural functions. J. Ethn. Foods 2017, 4, 28–35. [Google Scholar] [CrossRef]
- Akissoé, N.; Hounhouigan, J.; Mestres, C.; Nago, M. How blanching and drying affect the color and functional characteristics of yam (Dioscorea cayenensis-rotundata) flour. Food Chem. 2003, 82, 257–264. [Google Scholar] [CrossRef]
- Montañez, S.J.L.; Venegas, G.J.; Bernardno, N.A.; González, C.L.; Yañez, F.J. Chemical Characterization and Nutritional Evaluation of Mountain’s yam (Dioscorea remotiflora Kunth). Tubers 2014, 5, 153–160. [Google Scholar]
- Sosa, V.; Nova, J. Linajes de angiospermas endémicas en México: Zonas de alto endemismo para la conservación. Acta Botánica Mex. 2012, 100, 293–315. Available online: https://abm.ojs.inecol.mx/index.php/abm/article/view/38 (accessed on 13 January 2021). [CrossRef]
- Santacruz, R.F.; Casas, S.J.F.; Pérez, P.R.; Rodríguez, G.E.; Torres, M.M.I.; Castillo, H.C.; Iturbe, S.I. Conservación, manejo y aprovechamiento del camote de cerro (Dioscorea spp.) en el estado de Jalisco, México. Av. Investig. Científica CUCBA 2005, 16, 179–183. Available online: http://www.floradejalisco.cucba.udg.mx/sites/default/files/publicaciones1/avances/avances_2005/Agronomia/SantacruzRuvalcabaFernando/SantacruzRuvalcabaFernando.pdf (accessed on 8 January 2022).
- Miranda, A.G.; Soto, J.L.M.; Ruiz, I.G. Parcial caracterización de nuevos almidones obtenidos del tubérculo de camote del cerro (Dioscorea spp.). Rev. Iberoam. Tecnol. Postcosecha 2008, 9, 81–88. Available online: https://www.redalyc.org/pdf/813/81311226011.pdf (accessed on 16 March 2023).
- González, V.M.E. El ñame (Dioscorea spp.) Características, usos y valor medicinal. Aspectos de importancia en el desarrollo de su cultivo. Cultiv. Trop. 2012, 33, 5–15. Available online: http://scielo.sld.cu/scielo.php?pid=S0258-59362012000400001&script=sci_arttext&tlng=en (accessed on 24 September 2020).
- Yu, S.; Lan, X.; Zhou, J.; Gao, K.; Zhong, C.; Xie, J. Dioscorea composita WRKY3 positively regulates salt-stress tolerance in transgenic Arabidopsis thaliana. J. Plant Physiol. 2022, 269, 153592. [Google Scholar] [CrossRef]
- Velázquez, H.J.M.; Durán, P.N.; Ruíz, C.J.A.; González, E.D.R.; Santacruz, R.F.; Gallegos, R.A. Distribución geográfica y usos de especies del género Dioscorea: Geographic distribution and uses of species of the genus Dioscorea. e-CUCBA 2022, 19, 141–150. Available online: http://e-cucba.cucba.udg.mx/index.php/e-Cucba/article/view/273/263 (accessed on 15 September 2022). [CrossRef]
- Atwater, D.Z.; Ervine, C.; Barney, J.N. Climatic niche shifts are common in introduced plants. Nat. Ecol. Evol. 2018, 2, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Bernabe, A.A.; Santacruz, R.F.; Cruz, S.F. Effect of plant growth regulators on plant regeneration of Dioscorea remotiflora (Kunth) through nodal explants. Plant Growth Regul. 2012, 68, 293–301. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Lyons, J.B.; Chilaka, C.A. The Dioscorea Genus (Yam) An Appraisal of Nutritional and Therapeutic Potentials. Foods 2020, 9, 1304. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, H.J.M.; Ruíz, C.J.A.; Durán, P.N.; González, E.D.R.; Santacruz, R.F.; García, R.G.E.; Gallegos-Rodríguez, A. Eco-Geography of Dioscorea composita (Hemsl.) in México and Central America under the Influence of Climate Change. Sustainability 2023, 15, 12320. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Osorio, O.L.; Jiménez, G.D. An exhaustive analysis of heuristic methods for variable selection in ecological niche modeling and species distribution modeling. Ecol. Inform. 2019, 53, 100983. [Google Scholar] [CrossRef]
- Chandra, A.; Naithani, H.B.; Verma, P.K.; Saxena, J.; Prajapati, S. Plant diversity assessment of selected forest sites of Gaya district of Bihar, India. J. Appl. Nat. Sci. 2021, 13, 424–432. [Google Scholar] [CrossRef]
- Ghorbani, A.; Bahrami, B. The influence environmental factors on the distribution of plant species in the southeast rangelands of Sabalan. Watershed Manag. Res. J. 2017, 30, 15–29. [Google Scholar] [CrossRef]
- Sánchez, G.J.D.J.; Ruiz, C.J.A.; García, G.M.; Ojeda, G.R.; Larios, L.D.L.C.; Holland, J.B.; García, R.G.E. Ecogeography of teosinte. PLoS ONE 2018, 13, e0192676. [Google Scholar] [CrossRef]
- Aburto-Cansino, G.N.; Ruiz-Corral, J.A.; Sánchez González, J.D.J.; González Eguiarte, D.R. Temperaturas cardinales de desarrollo del teocintle (Zea spp.). Rev. Mex. Cienc. Agrícolas 2018, 9, 1269–1281. [Google Scholar] [CrossRef]
- Dormann, C.F.; McPherson, J.M.; Araujo, M.B.; Bivand, R.; Bolliger, J.; Carl, G. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Peterson, A.T.; Nakazawa, Y. Environmental data sets matter in ecological niche modelling: An example with Solenopsis invicta and Solenopsis richteri. Glob. Ecol. Biogeogr. 2008, 17, 135–144. [Google Scholar] [CrossRef]
- Feng, X.; Park, D.S.; Liang, Y.; Pandey, R.; Papeş, M. Collinearity in ecological niche modeling: Confusions and challenges. Ecol. Evol. 2019, 9, 10365–10376. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, R.; Gao, L.; Huang, D.; Fan, Y.; Liu, C.; Chen, J. Predicting the current and future distributions of Pennisetum alopecuroides (L.) in China under climate change based on the MaxEnt model. PLoS ONE 2023, 18, e0281254. [Google Scholar] [CrossRef]
- UNEP (United Nations Environmental Program). World Atlas of Desertification, 2nd ed.; Middleton, N., Thomas, D.S.G., Eds.; Arnold: London, UK, 1997; pp. 1–182. [Google Scholar]
- Rodríguez, W. Botánica, domesticación y fisiología del cultivo de ñame (Dioscorea alata). Agron. Mesoam. 2000, 11, 133–152. Available online: https://www.redalyc.org/pdf/437/43711221.pdf (accessed on 28 September 2022). [CrossRef]
- Avalo, D.M.R.; García, M.B. Producción y diversificación sostenible del cultivo de ñame (Dioscorea spp.) en condiciones de sequía agrícola en el municipio de Jiguaní. Agrisost 2021, 27, 1–7. Available online: https://revistas.reduc.edu.cu/index.php/agrisost/article/view/e10333-1 (accessed on 10 May 2022).
- Mueller, L.; Schindler, U.; Mirschel, W.; Shepherd, T.G.; Ball, B.C.; Helming, K.; Rogasik, J.; Eulenstein, F.; Wiggering, H. Assessing the productivity function of soils. A review. Sustain. Agric. 2010, 30, 601–614. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Geografía. INEGI Suelos; Instituto Nacional de Estadística y Geografía: Mexico City, Mexico, 2007; p. 36. Available online: https://apps1.semarnat.gob.mx:8443/dgeia/informe_12/pdf/Cap3_suelos.pdf (accessed on 18 May 2022).
- López, F.A.J.; Hernández, C.D. Cambio climático y agricultura: Una revisión de la literatura con énfasis en América Latina. El Trimest. Económico. 2016, 83, 459–496. [Google Scholar] [CrossRef]
- Ruiz, C.J.A.; Medina, G.G.; González, A.I.J.; Flores, L.H.E.; Ramírez, O.G.; Ortiz, T.C.; Byerly, M.K.F. Requerimientos Agroecológicos de Cultivos, 3rd ed.; INIFAP-Prometeo Editores: Jalisco, Mexico, 2013; p. 564. Available online: https://www.researchgate.net/publication/343047223_REQUERIMIENTOS_AGROECOLOGICOS_DE_CULTIVOS_2da_Edicion (accessed on 10 June 2022).
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen (Para Adaptarlo a las Condiciones de la República Mexicana), 5th ed.; Instituto de Geografía-UNAM: Mexico City, Mexico, 2004; p. 90. Available online: http://www.publicaciones.igg.unam.mx/index.php/ig/catalog/book/83 (accessed on 11 May 2022).
- Thompson, A.K.; Oduro, I. Yams: Botany, Production and Uses, 1st ed.; CABI: Boston, MA, USA, 2021; p. 296. [Google Scholar] [CrossRef]
- Patricio, V.E.G.; Rodrigo, A.R.J.; Isaías, M.A.R.; Cristina, L.V.I.; del Pilar, P.M.N.; Santiago, E.V.J. Inhibición de la brotación del tubérculo de papa: Una revisión de los métodos empleados Inhibition of potato tuber sprouting: A review of the methods employed. J. Selva Andin. Biosph. 2018, 6, 55–64. Available online: http://scielo.org.bo/pdf/jsab/v6n2/v6n2_a04.pdf (accessed on 10 August 2022).
- Coursey, D. Yams: Dioscorea spp., 2nd ed.; Wiley: London, UK, 1967; pp. 70–74. Available online: https://agris.fao.org/agrissearch/search.do?recordID=US201303063121 (accessed on 10 August 2022).
- Yang, J.; Huang, Y.; Jiang, X.; Chen, H.; Liu, M.; Wang, R. Potential geographical distribution of the endangered plant Isoetes under human activities using MaxEnt and GARP. Glob. Ecol. Conserv. 2022, 38, e02186. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley, G.M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MaxEnt ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Fois, M.A.; Cuena, L.G.; Fenu, G. Bacchetta Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef]
- Shen, L.; Xu, J.; Luo, L.; Hu, H.; Meng, X.; Li, X.; Chen, S. Predicting the potential global distribution of diosgenin-contained Dioscorea species. Chin. Med. 2018, 13, 58. [Google Scholar] [CrossRef]
- Viruel, J.; Catalan, P.; Segarr, M.J.G. Latitudinal environmental niches and riverine barriers shaped the phylogeography of the central Chilean endemic Dioscorea humilis (Dioscoreaceae). PLoS ONE 2014, 9, e110029. [Google Scholar] [CrossRef]
- Yin, D.S.; Chen, F.; Chen, Z.; Guan, W.F. Morpho- anatomical and physiological responses of two Dendranthema species to waterlogging. Environ. Exp. Botany 2010, 68, 122–130. [Google Scholar] [CrossRef]
- Jiménez, J.D.L.C.; Moreno, L.P.; Magnitskiy, S. Respuesta de las plantas a estrés por inundación. Una revisión. Revista Colombiana Ciencias Hortícolas 2012, 6, 96–109. Available online: https://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S2011-21732012000 100010 (accessed on 10 August 2022). [CrossRef]
- Medina, G.C.; Giménez de Azcárate, J.; Velázquez, M.A. Las comunidades vegetales del bosque de coníferas altimontano en el macizo del Tancítaro, Michoacán, México. Acta Botánica Mex. 2020, 127, 1584. [Google Scholar] [CrossRef]
- Schaal, B.A.; Hayworth, D.A.; Olsen, K.M.; Rauscher, J.T.; Smith, W.A. Phylogeographic studies in plants: Problems and perspectives. Mol. Ecol. 1998, 7, 465–474. Available online: https://www.researchgate.net/profile/Kenneth-Olsen-4/publication/227339790_Phylogeographic_studies_in_plants_Problems_and_prospects/links/5dfd016da6fdcc2837318eee/Phylogeographic-studies-in-plants-Problems-and-prospects.pdf (accessed on 28 August 2022). [CrossRef]
- Dillon, M.O.; Tu, T.; Xie, L.; Quipuscoa, S.V.; Wen, J. Biogeographic diversification in Nolana (Solanaceae), a ubiquitous member of the Atacama and Peruvian Deserts along the western coast of South America. J. Syst. Evol. 2009, 47, 457–476. [Google Scholar] [CrossRef]
- Schmidt, J.R.; Sytsma, K.J. Phylogenetics of Puya (Bromeliaceae): Placement, major lineages, and evolution of Chilean species. Am. J. Bot. 2010, 97, 337–356. [Google Scholar] [CrossRef]
- Gao, X.; Liu, J.; Huang, Z. The impact of climate change on the distribution of rare and endangered tree Firmiana kwangsiensis using the MaxEnt modeling. Ecol. Evol. 2022, 12, e9165. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Cao, Y.; DeWalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using MaxEnt to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecol. Model. 2013, 259, 30–39. [Google Scholar] [CrossRef]
- Lema De la Torre, L.E.; Rangel, J.A.M.; Carrera, J.A. Estudio etnobotánico del bosque protector cascada de peguche, otavalo, ecuador: Propuesta de estrategias de conservación. Recinatur Int. J. Appl. Sci. Nat. Tour. 2019, 1, 1–11. [Google Scholar]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on MaxEnt model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Ramírez, O.G.; Peralta, I.E.; Rodríguez, G.E.; Sahagún, C.J.; Chávez, S.J.L.; Medina, H.T.C.; Rodríguez, P.J.E. Edaphoclimatic Descriptors of Wild Tomato Species (Solanum Sect. Lycopersicon) and Closely Related Species (Solanum Sect. Juglandifolia and Sect. Lycopersicoides) in South America. Front. Genet. 2021, 12, 748979. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2010; Volume 1, p. 5. Available online: https://earthobservations.org/about_geo.shtml (accessed on 13 August 2022).
- O’brien, R.M. Caution Regarding Rules of Thumb for Variance Inflation Factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.2.1; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 August 2022).
- INEGI. Conjunto de Datos Vectoriales de Edafología Serie 4, Escala 1:250,000; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2017. [Google Scholar]
- Thouverai, E.; Marcantonio, M.; Lenoir, J.; Galfré, M.; Marchetto, E.; Bacaro, G.; Rocchini, D. Integrals of life: Tracking ecosystem spatial heterogeneity from space through the area under the curve of the parametric Rao’s Q index. Ecol. Complex. 2023, 52, 101029. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elithm, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J.A. Statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 171, 43–57. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in MaxEnt: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Mazzolari, A.C.; Millán, E.N.; Bringa, E.M.; Vázquez, D.P. Modeling habitat suitability and spread dynamics of two invasive rose species in protected areas of Mendoza, Argentina. Ecol. Complex. 2020, 44, 100868. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agroclimatic Region | Annual Moisture Availability Index | Annual Mean Temperature (°C) | Total Accessions |
|---|---|---|---|
| Semiarid very warm | 0.2–0.5 | >26 | 17 |
| Semiarid warm | 0.2–0.5 | 22–26 | 22 |
| Semiarid semiwarm | 0.2–0.5 | 18 a 22 | 14 |
| Semiarid temperate | 0.2–0.5 | 12 a 18 | 3 |
| Dry–subhumid very warm | 0.5–0.65 | > 26 | 20 |
| Dry–subhumid warm | 0.5–0.65 | 22–26 | 66 |
| Dry–subhumid semiwarm | 0.5–0.65 | 18–22 | 88 |
| Dry–subhumid temperate | 0.5–0.65 | 12–18 | 7 |
| Humid–subhumid very warm | 0.5–0.65 | >26 | 19 |
| Humid–subhumid warm | 0.65–1.0 | 22–26 | 67 |
| Humid–subhumid semiwarm | 0.65–1.0 | 18–22 | 98 |
| Humid–subhumid temperate | 0.65–1.0 | 12–18 | 11 |
| Humid very warm | >1.0 | >26 | 2 |
| Humid warm | >1.0 | 22–26 | 26 |
| Humid semiwarm | >1.0 | 18–22 | 12 |
| Humid temperate | >1.0 | 12–18 | 6 |
| Humid semicold | >1.0 | 5–12 | 3 |
| FAO Soil Unit | Soil Texture | Total Accessions |
|---|---|---|
| Lithosol | Coarse | 108 |
| Regosol calcaric | Coarse | 57 |
| Regosol eutric | Coarse | 209 |
| Faozem haplic | Coarse | 34 |
| Vertisol cromic | Fine | 39 |
| Solonchak ortic | Fine | 22 |
| Fluvisol eutric | Medium | 10 |
| Fluvisol calcaric | Coarse | 1 |
| Environmental Variables | Min | Max | Optimum |
|---|---|---|---|
| 1. Precipitation of the warmest quarter (mm) | 240 | 1204 | 400–884 |
| 2. Precipitation of the driest month (mm) | 1 | 73 | 1–7 |
| 3. Annual mean precipitation (mm) | 444 | 2886 | 700–1299 |
| 4. May–October mean precipitation (mm) | 344 | 1943 | 700–1199 |
| 5. November–April mean precipitation | 23 | 863 | 30–100 |
| 6. Annual moisture availability index | 0.27 | 2.32 | 0.40–0.99 |
| 7. November–April availability index | 0.026 | 1.83 | 0.030–1300 |
| 8. May–October availability index | 0.005 | 1.47 | 0.009–1.4 |
| 9. Maximum maximorum temperature (°C) | 24.61 | 41.17 | 29–37 |
| 10. Minimum minimorum temperature (°C) | 1.7 | 18.2 | 5–15 |
| 11. Annual mean temperature (°C) | 14.66 | 28.51 | 19–27 |
| 12. May–October mean temperature | 9.13 | 29.88 | 19–26 |
| 13. November–April mean temperature | 7.95 | 27.83 | 19–26 |
| 14. Annual thermal oscillation (°C) | 10.42 | 19.54 | 13.16 |
| 15. Annual temperature range (°C) | 1.54 | 14.52 | 3–7 |
| 16. Soil texture | Sandy | Fine | Medium |
| 17. May–October mean photoperiod (h) | 12.5 | 12.9 | 12.6–12.9 |
| 18. November–April mean photoperiod (h) | 10.97 | 11.47 | 11.10–11.39 |
| 19. Growing season | 120–190 | ||
| 20. Altitude (mm) | 6 | 4295 | 200–1800 |
| Environmental Variables | Contribution (%) | Permutation Importance (%) |
|---|---|---|
| Precipitation of the warmest quarter (mm) | 42.4 | 49 |
| Precipitation of the driest month (mm) | 17.5 | 2.8 |
| Minimum temperature of the coldest month (°C) | 15 | 31 |
| November–April mean solar radiation (w/m2) | 10 | 0.8 |
| Annual mean relative humidity (%) | 8.5 | 3.6 |
| Annual moisture availability index | 5.7 | 7.4 |
| May–October mean temperature (°C) | 0.9 | 5.3 |
| Parameter | Value |
|---|---|
| AUC of the ROC curve | 0.935 |
| Mean AUC ratio | 1.679 |
| Omission rate (%) | 0.5 |
| AICc | 12,592.861 |
| Delta AICc | 0 |
| W AICc | 0.9999 |
| Optimum regularization multiplier | 3.0 |
| Institution/Source | Institution/Department | Accessions |
|---|---|---|
| Universidad Nacional Autónoma de México (UNAM). | Instituto de Biología | 169 |
| Instituto de Ecología (INECOL). | Xalapa Veracruz | 30 |
| Universidad Autónoma de Querétaro (UAQ) | Facultad de Ciencias Naturales | 3 |
| Instituto Nacional de Estadística y Geografía (INEGI). | Departamento de Botánica | 2 |
| Universidad Autónoma de Aguascalientes (UAA). | Centro de Ciencias Básicas | 2 |
| Universidad Autónoma de Veracruz (UPAV) (CIB). | Instituto de Investigaciones Biológicas | 1 |
| Universidad Autónoma de San Luis Potosí (UASLP). | Instituto de Investigación de Zonas Desérticas | 3 |
| Colegio de la Frontera Sur (ECO SUR). | Herbario San Cristóbal | 3 |
| Universidad de Guadalajara (CUCBA, CUC SUR). | Herbario IBUG, Herbario ZEA | 6 |
| Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán. | Herbario Facultad de Biología Universidad Michoacana | 6 |
| Artículos científicos/Inventarios florísticos de los estados de Oaxaca, Chiapas, Veracruz, Tabasco, Guerrero, Puebla, Jstor Plant Science. | 20 | |
| Universidad Autónoma de Nuevo León (UNL). | Facultad de Ciencias Biológicas | 1 |
| La Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). | Herbario digital de CONABIO | 3 |
| Trópicos.org. | 3 | |
| Red de Herbarios del Noroeste de México. | 13 | |
| GBIF | 215 | |
| Total | 480 |
| Variable | Description | Temporal Scale |
|---|---|---|
| BIO01 | Annual mean temperature | Annual |
| BIO02 | Mean diurnal range | Variation |
| BIO03 | Isothermality | Variation |
| BIO04 | Temperature seasonality | Variation |
| BIO05 | Maximum temperature of the warmest month | Month |
| BIO06 | Minimum temperature of the coldest month | Month |
| BIO07 | Temperature annual range | Annual |
| BIO08 | Mean temperature of the wettest quarter | Quarter |
| BIO09 | Mean temperature of the driest quarter | Quarter |
| BIO10 | Mean temperature of the warmest quarter | Quarter |
| BIO11 | Mean temperature of the coldest quarter | Quarter |
| BIO12 | Annual precipitation | Annual |
| BIO13 | Precipitation of the wettest month | Month |
| BIO14 | Precipitation of the driest month | Month |
| BIO15 | Precipitation seasonality | Variation |
| BIO16 | Precipitation of the wettest quarter | Quarter |
| BIO17 | Precipitation of the driest quarter | Quarter |
| BIO18 | Precipitation of the warmest quarter | Quarter |
| BIO19 | Precipitation of the coldest quarter | Quarter |
| N-AMT | November–April mean temperature | Seasonal |
| M-OMT | May–October mean temperature | Seasonal |
| M-OXT | Maximum temperature May–October | Seasonal |
| N-AXT | November–April maximum temperature | Seasonal |
| AXT | Annual maximum temperature | Annual |
| M-OIT | May–October minimum temperature | Seasonal |
| N-AIT | November–April minimum temperature | Seasonal |
| AIT | Annual minimum temperature | Annual |
| ATO | Annual thermal oscillation | Annual |
| M-OP | May–October precipitation | Seasonal |
| N-AP | November–April precipitation | Seasonal |
| M-OPH | May–October photoperiod | Seasonal |
| N-APH | November–April photoperiod | Seasonal |
| AMI | Annual moisture index | Annual |
| M-OMI | May–October mean moisture index | Seasonal |
| N-AMI | November–April mean moisture index | Seasonal |
| ASR | Annual mean solar radiation | Annual |
| M-OSR | May–October mean solar radiation | Seasonal |
| N-ASR | November–April solar radiation | Seasonal |
| ARH | Annual relative humidity | Annual |
| M-ORH | May–October relative humidity | Seasonal |
| N-ARH | November–April relative humidity | Seasonal |
| GSL | Growing season length | Seasonal |
| Criteria |
|---|
| All candidate models |
| Statistically significant models |
| Models meeting omission rate criteria |
| Models meeting AICc criteria |
| Models meeting high AUC value |
| Statistically significant models meeting omission rate criteria |
| Statistically significant models meeting AICc criteria |
| Statistically significant models meeting high AUC value |
| Statistically significant models meeting omission rate criteria, AICc criteria, and AUC criteria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velázquez-Hernández, J.M.; Ruíz-Corral, J.A.; Durán-Puga, N.; Macías, M.Á.; González-Eguiarte, D.R.; Santacruz-Ruvalcaba, F.; García-Romero, G.E.; Gallegos-Rodríguez, A. Ecogeography of Dioscorea remotiflora Kunth: An Endemic Species from Mexico. Plants 2023, 12, 3654. https://doi.org/10.3390/plants12203654
Velázquez-Hernández JM, Ruíz-Corral JA, Durán-Puga N, Macías MÁ, González-Eguiarte DR, Santacruz-Ruvalcaba F, García-Romero GE, Gallegos-Rodríguez A. Ecogeography of Dioscorea remotiflora Kunth: An Endemic Species from Mexico. Plants. 2023; 12(20):3654. https://doi.org/10.3390/plants12203654
Chicago/Turabian StyleVelázquez-Hernández, Jocelyn Maira, José Ariel Ruíz-Corral, Noé Durán-Puga, Miguel Ángel Macías, Diego Raymundo González-Eguiarte, Fernando Santacruz-Ruvalcaba, Giovanni Emmanuel García-Romero, and Agustín Gallegos-Rodríguez. 2023. "Ecogeography of Dioscorea remotiflora Kunth: An Endemic Species from Mexico" Plants 12, no. 20: 3654. https://doi.org/10.3390/plants12203654