
Environmental Filtering and Dispersal Limitations Driving the Beta Diversity Patterns at Different Scales of Secondary Evergreen Broadleaved Forests in the Suburbs of Hangzhou

Abstract
:1. Introduction
2. Results
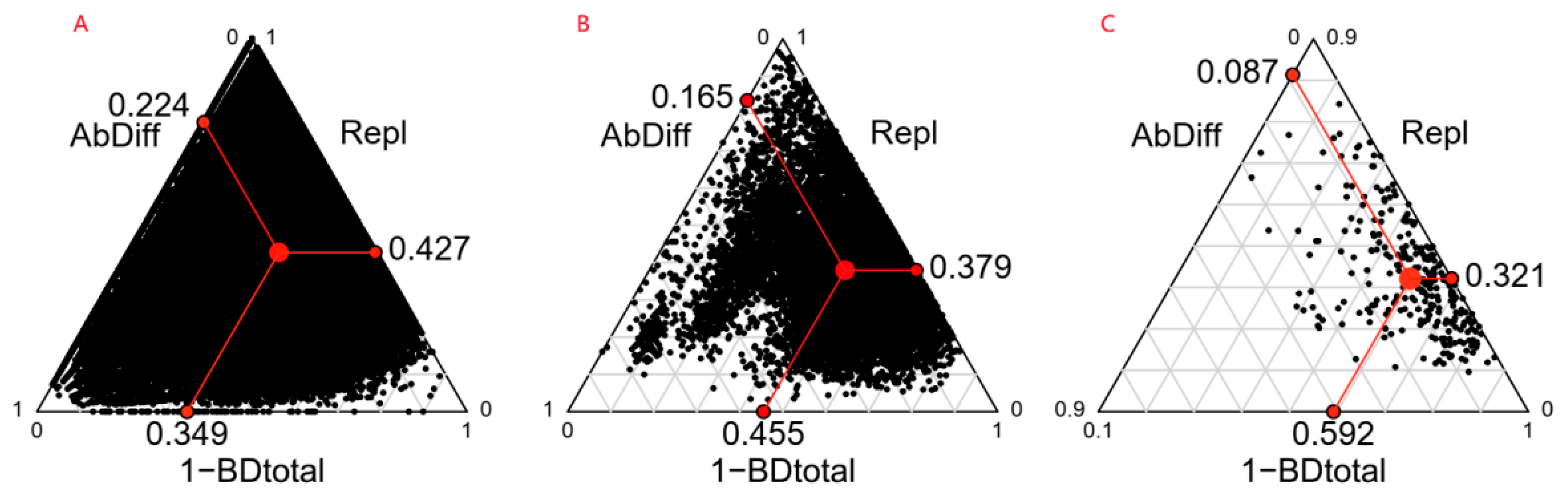
2.1. Community Beta Diversity and Its Components
2.2. Variance Decomposition of Community Beta Diversity
3. Discussion
3.1. The Changing Pattern of Beta Diversity of a Suburban Secondary Evergreen Broadleaved Forest Community
3.2. Community Construction Mechanism
4. Materials and Methods
4.1. Description of the Study Site
4.2. Construction and Investigation of the Dynamic Monitoring Sample Plots
4.3. Data Preparation and Processing
4.4. Analysis of Beta Diversity Patterns and Partitioning Beta Diversity
4.5. Investigation of Driving Forces
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Abundance | Importance Values |
|---|---|---|
| Schima superba | 1745 | 17.9% |
| Camellia fraterna | 6396 | 7.8% |
| Symplocos anomala | 5677 | 7.2% |
| Cyclobalanopsis glauca | 2117 | 6.0% |
| Eurya rubiginosa | 4151 | 5.3% |
| Cunninghamia lanceolata | 679 | 4.3% |
| Rhododendron ovatum | 2717 | 4.1% |
| Osmanthus cooperi | 1394 | 3.3% |
| Loropetalum chinense | 1197 | 2.7% |
| Castanopsis sclerophylla | 325 | 2.7% |
| Symplocos stellaris | 1336 | 2.7% |
| Quercus serrata | 332 | 2.5% |
| Lithocarpus glaber | 440 | 2.4% |
| Cleyera japonica | 994 | 2.2% |
| Number of Plots | Number of Species | Elevation | Soil Organic Matter (g/kg) | Soil Total N Content (g/kg) | Soil Total P Content (g/kg) | Soil Available K (mg/kg) | Soil Available N (mg/kg) | Soil Available P (mg/kg) |
|---|---|---|---|---|---|---|---|---|
| 150 (20 m × 20 m) | 93 | 390 ± 42 | 118 ± 54 | 3.8 ± 1.5 | 0.35 ± 0.21 | 98 ± 33 | 323 ± 97 | 3.7 ± 1.8 |

References
- Zhang, L.; Su, Z.; Chen, B. Interspecific Relationships in the Forest Community Dominated by Pinus Kwangtungensis, an Endangered Species Endemic to China. Front. For. China 2007, 2, 128–135. [Google Scholar] [CrossRef]
- Duchesneau, R.; Lesage, I.; Messier, C.; Morin, H. Effects of Light and Intraspecific Competition on Growth and Crown Morphology of Two Size Classes of Understory Balsam Fir Saplings. Ecol. Manag. 2001, 140, 215–225. [Google Scholar] [CrossRef]
- Mi, X.; Ma, K.; Zhu, Y. A mechanism of plant species coexistence: The negative density-dependent hypothesis. Biodivers. Sci. 2009, 17, 594. [Google Scholar] [CrossRef]
- Bai, S.; Zhou, G.; Wang, Y.; Liang, Q.; Chen, J.; Cheng, Y.; Shen, R. Plant Species Diversity and Dynamics in Forests Invaded by Moso Bamboo (Phyllostachys Edulis) in Tianmu Mountain Nature Reserve. Biodivers. Sci. 2013, 21, 288–295. [Google Scholar] [CrossRef]
- He, J.; Lin, S.; Kong, F.; Yu, J.; Zhu, H.; Jiang, H. Determinants of the Beta Diversity of Tree Species in Tropical Forests: Implications for Biodiversity Conservation. Sci. Total Environ. 2020, 704, 135301. [Google Scholar] [CrossRef]
- Stevenson, P.R.; Aldana, A.M.; Cárdenas, S.; Negret, P.J. Flooding and Soil Composition Determine Beta Diversity of Lowland Forests in Northern South America. Biotropica 2018, 50, 568–577. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Dolos, K.; Reineking, B.; Beierkuhnlein, C. Current Measures for Distance Decay in Similarity of Species Composition Are Influenced by Study Extent and Grain Size. Glob. Ecol. Biogeogr. 2012, 21, 1203–1212. [Google Scholar] [CrossRef]
- Scudeller, V.V.; Vegas-Vilarrúbia, T. Distribution and β-Diversity of Tree Species in Igapó Forests (Negro River Basin, Brazilian Amazon). J. Veg. Sci. 2018, 29, 1052–1064. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, J.; Xu, S.; Lei, Y.; Yang, R.; Shi, L.; Wang, X.; Huang, Z. Bacterioplankton Community Variation in Bohai Bay (China) Is Explained by Joint Effects of Environmental and Spatial Factors. Microbiologyopen 2020, 9, e997. [Google Scholar] [CrossRef]
- Bini, L.M.; Landeiro, V.L.; Padial, A.A.; Siqueira, T.; Heino, J. Nutrient Enrichment Is Related to Two Facets of Beta Diversity for Stream Invertebrates across the United States. Ecology 2014, 95, 1569–1578. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A Trait-Based Test for Habitat Filtering: Convex Hull Volume. Ecology 2006, 87, 1465–1471. [Google Scholar]
- Swenson, N.G. The Assembly of Tropical Tree Communities—The Advances and Shortcomings of Phylogenetic and Functional Trait Analyses. Ecography 2013, 36, 264–276. [Google Scholar] [CrossRef]
- Muscarella, R.; Uriarte, M.; Erickson, D.L.; Swenson, N.G.; Kress, W.J.; Zimmerman, J.K. Variation of Tropical Forest Assembly Processes across Regional Environmental Gradients. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 52–62. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Avois-Jacquet, C.; Tuomisto, H. Dissecting the Spatial Structure of Ecological Data at Multiple Scales. Ecology 2004, 85, 1826–1832. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed]
- De Cáceres, M.; Legendre, P.; Valencia, R.; Cao, M.; Chang, L.-W.; Chuyong, G.; Condit, R.; Hao, Z.; Hsieh, C.F.; Hubbell, S.; et al. The Variation of Tree Beta Diversity across a Global Network of Forest Plots. Glob. Ecol. Biogeogr. 2012, 21, 1191–1202. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the Importance of Ecological Niches from Stochastic Processes across Scales. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef]
- Medina, R.F.; Szendrei, Z.; Harrison, K.; Isaacs, R.; Averill, A.; Malo, E.A.; Rodriguez-Saona, C. Exploring Host-Associated Differentiation in the North American Native Cranberry Fruitworm, Acrobasis Vaccinii, from Blueberries and Cranberries. Entomol. Exp. Appl. 2014, 150, 136–148. [Google Scholar] [CrossRef]
- Jones, M.M.; Tuomisto, H.; Borcard, D.; Legendre, P.; Clark, D.B.; Olivas, P.C. Explaining Variation in Tropical Plant Community Composition: Influence of Environmental and Spatial Data Quality. Oecologia 2008, 155, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Gravel, D.; Canham, C.D.; Beaudet, M.; Messier, C. Reconciling Niche and Neutrality: The Continuum Hypothesis. Ecol. Lett. 2006, 9, 399–409. [Google Scholar] [CrossRef]
- Yao, L.; Wang, Z.; Zhan, X.; Wu, W.; Jiang, B.; Jiao, J.; Yuan, W.; Zhu, J.; Ding, Y.; Li, T.; et al. Assessment of Species Composition and Community Structure of the Suburban Forest in Hangzhou, Eastern China. Sustainability 2022, 14, 4304. [Google Scholar] [CrossRef]
- Jiao, J.; Wu, C.; Jiang, B.; Wang, Z.; Yuan, W.; Zhu, J.; Li, T.; Yang, S.; Yao, L. Negative Density Restricts the Coexistence and Spatial Distribution of Dominant Species in Subtropical Evergreen Broad-Leaved Forests in China. Forests 2022, 13, 1227. [Google Scholar] [CrossRef]
- Podani, J.; Schmera, D. Once Again on the Components of Pairwise Beta Diversity. Ecol. Inf. 2016, 32, 63–68. [Google Scholar] [CrossRef]
- Stuart, C.T.; Brault, S.; Rowe, G.T.; Wei, C.; Wagstaff, M.; McClain, C.R.; Rex, M.A. Nestedness and Species Replacement along Bathymetric Gradients in the Deep Sea Reflect Productivity: A Test with Polychaete Assemblages in the Oligotrophic North-west Gulf of Mexico. J. Biogeogr. 2017, 44, 548–555. [Google Scholar] [CrossRef]
- Guo, Y.; Xiang, W.; Wang, B.; Li, D.; Mallik, A.U.; Chen, H.Y.H.; Huang, F.; Ding, T.; Wen, S.; Lu, S.; et al. Partitioning Beta Diversity in a Tropical Karst Seasonal Rainforest in Southern China. Sci. Rep. 2018, 8, 17408. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.-F.; He, F. Partitioning Beta Diversity in a Subtropical Broad-leaved Forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Sfair, J.C.; Arroyo-Rodríguez, V.; Santos, B.A.; Tabarelli, M. Taxonomic and Functional Divergence of Tree Assemblages in a Fragmented Tropical Forest. Ecol. Appl. 2016, 26, 1816–1826. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community Assembly, Coexistence and the Environmental Filtering Metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, B.; Li, D.; Mallik, A.U.; Xiang, W.; Ding, T.; Wen, S.; Lu, S.; Huang, F.; He, Y.; et al. Effects of Topography and Spatial Processes on Structuring Tree Species Composition in a Diverse Heterogeneous Tropical Karst Seasonal Rainforest. Flora 2017, 231, 21–28. [Google Scholar] [CrossRef]
- Jin, C.; Jiang, B.; Ding, Y.; Yang, S.; Xu, Y.; Jiao, J.; Huang, J.; Yuan, W.; Wu, C. Functional Traits Change but Species Diversity Is Not Influenced by Edge Effects in an Urban Forest of Eastern China. Urban. Urban. Green. 2021, 64, 127245. [Google Scholar] [CrossRef]
- Guo, Y.; Li, D.; Wang, B.; Bai, K.; Xiang, W.; Li, X. C, N and P Stoichiometric Characteristics of Soil and Litter Fall for Six Common Tree Species in a Northern Tropical Karst Seasonal Rainforest in Nonggang, Guangxi, Southern China. Biodivers. Sci. 2017, 25, 1085–1094. [Google Scholar] [CrossRef]
- Baselga, A.; Bonthoux, S.; Balent, G. Temporal Beta Diversity of Bird Assemblages in Agricultural Landscapes: Land Cover Change vs. Stochastic Processes. PLoS ONE 2015, 10, e0127913. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning Abundance-based Multiple-site Dissimilarity into Components: Balanced Variation in Abundance and Abundance Gradients. Methods Ecol. Evol. 2017, 8, 799–808. [Google Scholar] [CrossRef]
- Chen, L.; Swenson, N.G.; Ji, N.; Mi, X.; Ren, H.; Guo, L.; Ma, K. Differential Soil Fungus Accumulation and Density Dependence of Trees in a Subtropical Forest. Science 2019, 366, 124–128. [Google Scholar] [CrossRef]
- Zhong, Y.; Chu, C.; Myers, J.A.; Gilbert, G.S.; Lutz, J.A.; Stillhard, J.; Zhu, K.; Thompson, J.; Baltzer, J.L.; He, F.; et al. Arbuscular Mycorrhizal Trees Influence the Latitudinal Beta-Diversity Gradient of Tree Communities in Forests Worldwide. Nat. Commun. 2021, 12, 3137. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Joseph, W.S.; Abu, S.K.; Almeyda, Z.A.M.; Alonso, A.; Baltzer, J.L.; et al. CTFS-ForestGEO: A worldwide network monitoring forests in an era of global change. Glob. Change Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef] [PubMed]



| Scale | Number of Samples | (a) | (b) | (c) | (a + b) | (b + c) | (d) |
|---|---|---|---|---|---|---|---|
| 10 m × 10 m | 600 | 0.0093 | 0.2796 | 0.3890 | 0.2889 | 0.6686 | 0.3221 |
| 20 m × 20 m | 150 | 0.0131 | 0.3932 | 0.2744 | 0.4063 | 0.6676 | 0.3193 |
| 50 m × 50 m | 24 | 0.1147 | 0.5011 | 0.1723 | 0.6158 | 0.6734 | 0.2119 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, L.; Jiang, B.; Jiao, J.; Wu, C. Environmental Filtering and Dispersal Limitations Driving the Beta Diversity Patterns at Different Scales of Secondary Evergreen Broadleaved Forests in the Suburbs of Hangzhou. Plants 2023, 12, 3057. https://doi.org/10.3390/plants12173057
Yao L, Jiang B, Jiao J, Wu C. Environmental Filtering and Dispersal Limitations Driving the Beta Diversity Patterns at Different Scales of Secondary Evergreen Broadleaved Forests in the Suburbs of Hangzhou. Plants. 2023; 12(17):3057. https://doi.org/10.3390/plants12173057
Chicago/Turabian StyleYao, Liangjin, Bo Jiang, Jiejie Jiao, and Chuping Wu. 2023. "Environmental Filtering and Dispersal Limitations Driving the Beta Diversity Patterns at Different Scales of Secondary Evergreen Broadleaved Forests in the Suburbs of Hangzhou" Plants 12, no. 17: 3057. https://doi.org/10.3390/plants12173057