
Identification of microRNAs from Medicinal Plant Murraya koenigii by High-Throughput Sequencing and Their Functional Implications in Secondary Metabolite Biosynthesis

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Sequence Analysis of M. koenigii Small RNAs
2.2. Identification of Conserved and Novel miRNAs in M. koenigii
2.3. Target Prediction of Conserved and Novel M. koenigii miRNAs and Their Functional Analysis
2.4. Human Target Gene Prediction of M. koenigii miRNAs
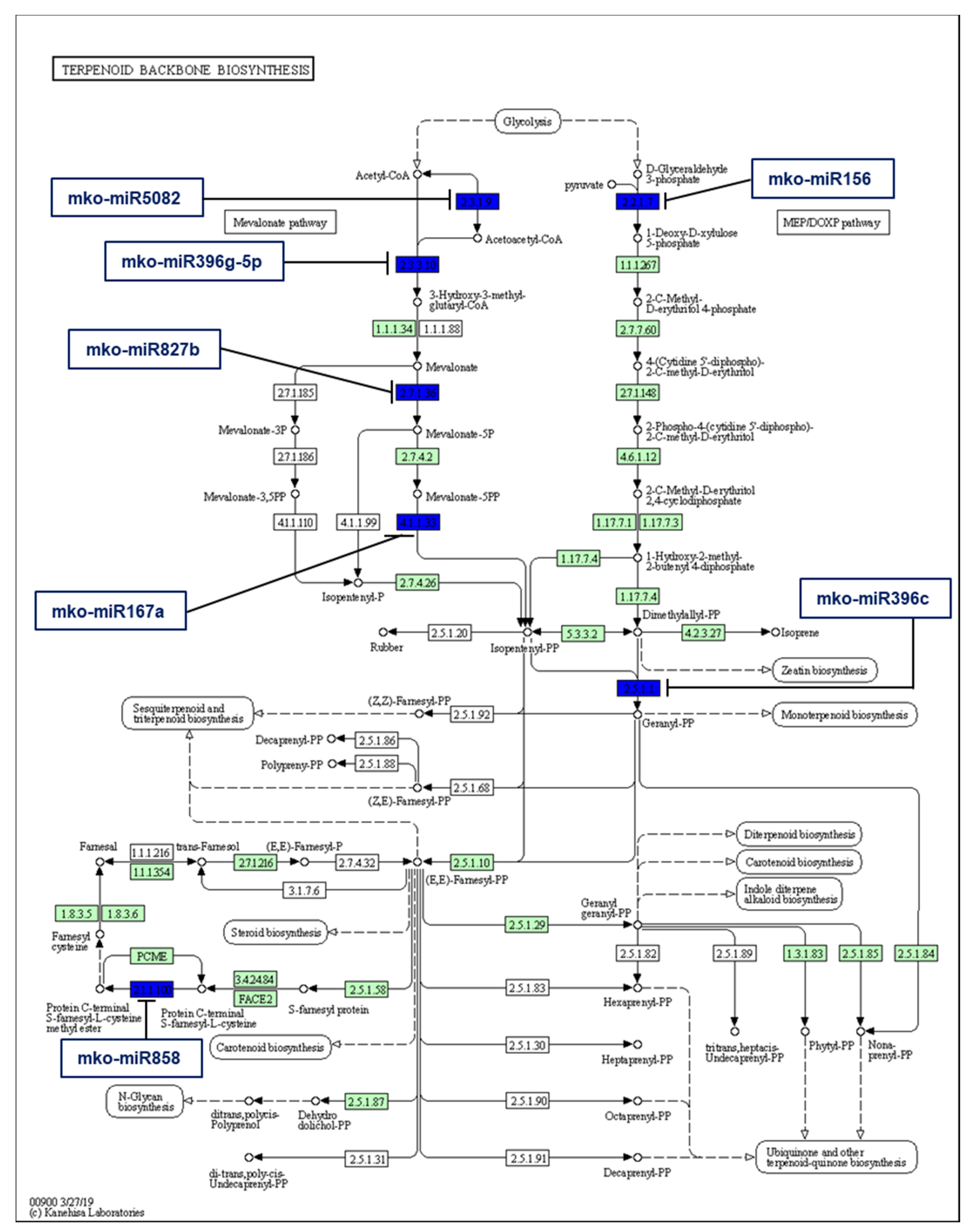
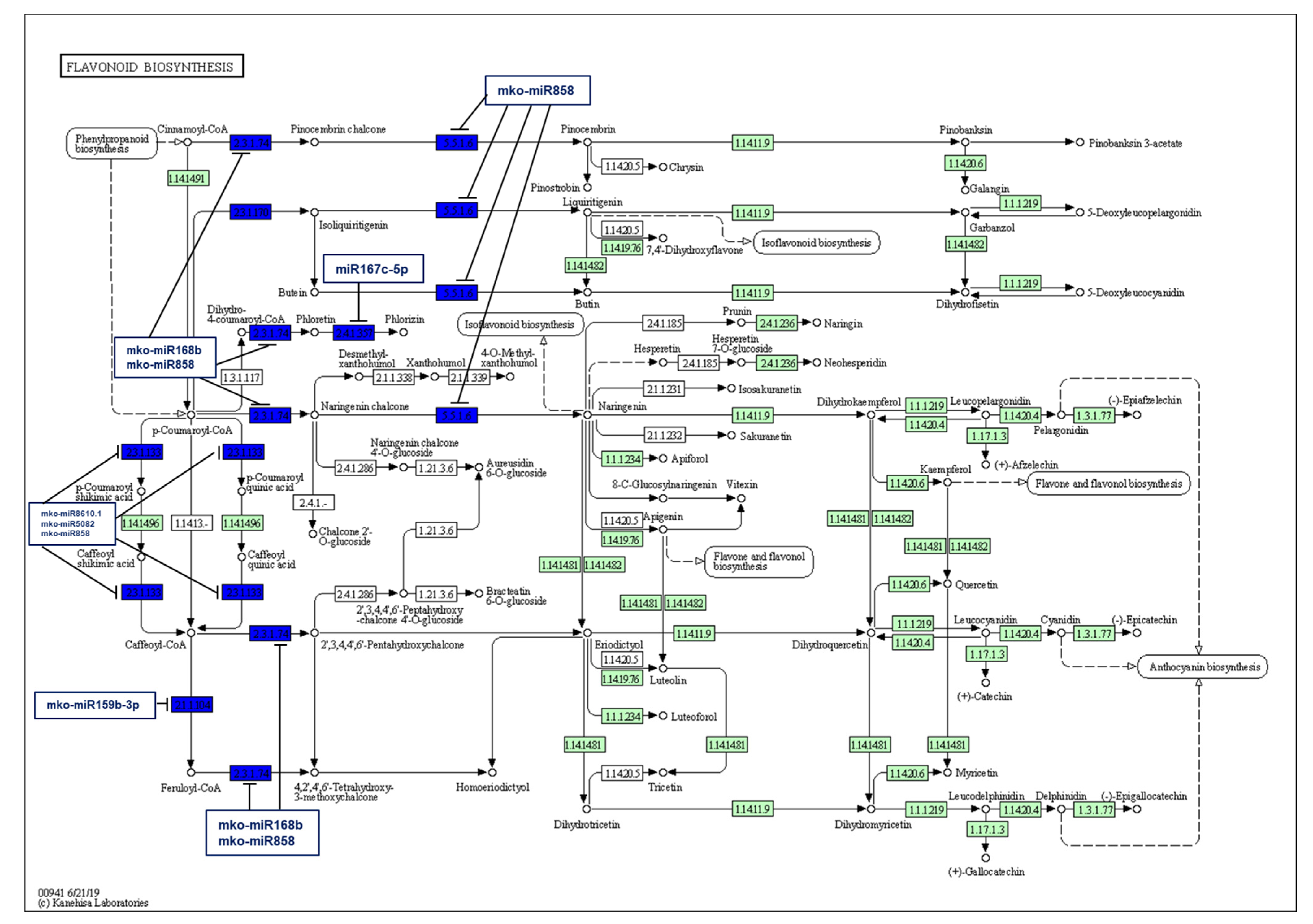
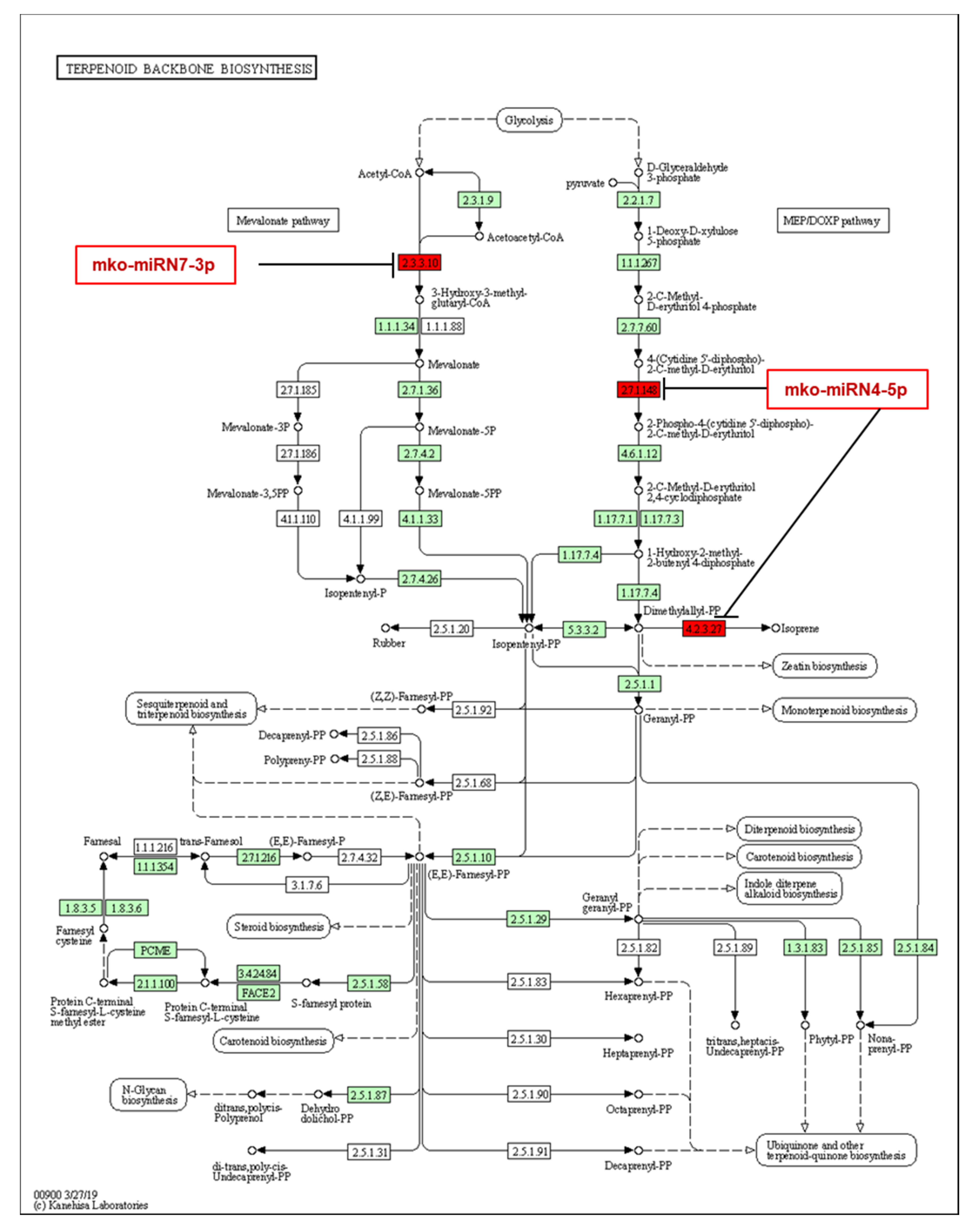
2.5. Identification of M. koenigii miRNA Targets Involved in Plant Secondary Metabolite Biosynthesis
2.6. Experimental Validation of M. koenigii miRNAs by qPCR
3. Discussion
4. Materials and Methods
4.1. Plant Materials and RNA Extraction
4.2. Small RNA Library Construction and Sequencing
4.3. Small RNA Sequencing Data Analysis
4.4. Prediction of M. koenigii miRNA Targets, Their Functional Annotation, and Pathway Analysis
4.5. Extraction of Small RNA and Experimental Validation of M. koenigii miRNAs by qPCR
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chauhan, B.; Dedania, J.; Mashru, D.R.C. Review on Murraya Koenigii: Versatile Role in Management of Human Health. World J. Pharm. Pharm. Sci. 2017, 6, 476–493. [Google Scholar] [CrossRef]
- Nandan, S.; Singh, S.K.; Singh, P.; Bajpai, V.; Mishra, A.K.; Joshi, T.; Mahar, R.; Shukla, S.K.; Mishra, D.K.; Kanojiya, S. Quantitative Analysis of Bioactive Carbazole Alkaloids in Murraya koenigii (L.) from Six Different Climatic Zones of India Using UPLC/MS/MS and Their Principal Component Analysis. Chem. Biodivers. 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P. A review on therapeutic use of Murraya koenigii (L.) Spreng. Int. Res. J. Mod. Eng. Technol. Sci. 2020, 2, 93–97. [Google Scholar]
- Balakrishnan, R.; Vijayraja, D.; Jo, S.H.; Ganesan, P.; Su-kim, I.; Choi, D.K. Medicinal profile, phytochemistry, and pharmacological activities of Murraya koenigii and its primary bioactive compounds. Antioxidants 2020, 9, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, A.; Sikha, M.S.; Ramesh, K.; Venkatesh, P.; Godugu, C. Modulation of cerulein-induced pancreatic inflammation by hydroalcoholic extract of curry leaf (Murraya koenigii). Phyther. Res. 2019, 33, 1510–1525. [Google Scholar] [CrossRef]
- Mahipal, P.; Pawar, R.S. Nephroprotective effect of Murraya koenigii on cyclophosphamide induced nephrotoxicity in rats. Asian Pac. J. Trop. Med. 2017, 10, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Husna, F.; Suyatna, F.D.; Arozal, W.; Poerwaningsih, E.H. Anti-Diabetic Potential of Murraya Koenigii (L.) and its Antioxidant Capacity in Nicotinamide-Streptozotocin Induced Diabetic Rats. Drug Res. 2018, 68, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.K.; Kandimalla, R.; Gogoi, B.; Dutta, K.N.; Choudhury, P.; Deb, P.K.; Devi, R.; Pal, B.C.; Talukdar, N.C. Phytochemical portfolio and anticancer activity of Murraya koenigii and its primary active component, mahanine. Pharmacol. Res. 2018, 129, 227–236. [Google Scholar] [CrossRef]
- Nalli, Y.; Khajuria, V.; Gupta, S.; Arora, P.; Riyaz-Ul-Hassan, S.; Ahmed, Z.; Ali, A. Four new carbazole alkaloids from Murraya koenigii that display anti-inflammatory and anti-microbial activities. Org. Biomol. Chem. 2016, 14, 3322–3332. [Google Scholar] [CrossRef]
- Chen, M.; Yin, X.; Lu, C.; Chen, X.; Ba, H.; Cai, J.; Sun, J. Mahanine induces apoptosis, cell cycle arrest, inhibition of cell migration, invasion and PI3K/AKT/mTOR signalling pathway in glioma cells and inhibits tumor growth in vivo. Chem. Biol. Interact. 2019, 299, 1–7. [Google Scholar] [CrossRef]
- Gupta, O.P.; Karkute, S.G.; Banerjee, S.; Meena, N.L.; Dahuja, A. Contemporary understanding of miRNA-based regulation of secondary metabolites biosynthesis in plants. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Reyes-Pérez, P.; Angulo-Bejarano, P.I.; Srivastava, A.; Ramalingam, S.; Sharma, A. Characterization of microRNAs from neem (Azadirachta indica) and their tissue-specific expression study in leaves and stem. 3 Biotech 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Sharma, A.; Ruiz-Manriquez, L.M.; Serrano-Cano, F.I.; Reyes-Pérez, P.R.; Alfaro, C.K.T.; Andrade, Y.E.B.; Aros, A.K.H.; Srivastava, A.; Paul, S. Identification of microRNAs and their expression in leaf tissues of guava (Psidium guajava L.) under salinity stress. Agronomy 2020, 10, 1920. [Google Scholar] [CrossRef]
- Stepien, A.; Knop, K.; Dolata, J.; Taube, M.; Bajczyk, M.; Barciszewska-Pacak, M.; Pacak, A.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Posttranscriptional coordination of splicing and miRNA biogenesis in plants. Wiley Interdiscip. Rev. RNA 2017, 8, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Li, Y.; Cao, X.; Qi, Y. MicroRNAs and Their Regulatory Roles in Plant-Environment Interactions. Annu. Rev. Plant Biol. 2019, 70, 489–525. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bravo Vázquez, L.A.; Márquez Nafarrate, M.; Gutiérrez Reséndiz, A.I.; Srivastava, A.; Sharma, A. The regulatory activities of microRNAs in non-vascular plants: A mini review. Planta 2021, 254, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Naya, L.; Paul, S.; Valdés-López, O.; Mendoza-Soto, A.B.; Nova-Franco, B.; Sosa-Valencia, G.; Reyes, J.L.; Hernández, G. Regulation of copper homeostasis and biotic interactions by microRNA 398b in common bean. PLoS ONE 2014, 9, e84416. [Google Scholar] [CrossRef]
- Paul, S. Identification and characterization of microRNAs and their targets in high-altitude stress-adaptive plant maca (Lepidium meyenii Walp). 3 Biotech 2017, 7, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative Regulation of Anthocyanin Biosynthesis in Arabidopsis by a miR156-Targeted SPL Transcription Factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef] [Green Version]
- Vashisht, I.; Mishra, P.; Pal, T.; Chanumolu, S.; Singh, T.R.; Chauhan, R.S. Mining NGS transcriptomes for miRNAs and dissecting their role in regulating growth, development, and secondary metabolites production in different organs of a medicinal herb, Picrorhiza kurroa. Planta 2015, 241, 1255–1268. [Google Scholar] [CrossRef]
- Li, J.Z.; Xie, M.Q.; Mo, D.; Zhao, X.F.; Yu, S.Y.; Liu, L.J.; Wu, C.; Yang, Y. Picroside II protects myocardium from ischemia/reperfusion–induced injury through inhibition of the inflammatory response. Exp. Ther. Med. 2016, 12, 3507–3514. [Google Scholar] [CrossRef] [Green Version]
- Boke, H.; Ozhuner, E.; Turktas, M.; Parmaksiz, I.; Ozcan, S.; Unver, T. Regulation of the alkaloid biosynthesis by miRNA in opium poppy. Plant Biotechnol. J. 2015, 13, 409–420. [Google Scholar] [CrossRef]
- Srivastava, S.; Singh, R.; Srivastava, G.; Sharma, A. Comparative Study of Withanolide Biosynthesis-Related miRNAs in Root and Leaf Tissues of Withania somnifera. Appl. Biochem. Biotechnol. 2018, 185, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Gupta, O.P.; Dahuja, A.; Sachdev, A.; Kumari, S.; Jain, P.K.; Vinutha, T.; Praveen, S. Conserved miRNAs modulate the expression of potential transcription factors of isoflavonoid biosynthetic pathway in soybean seeds. Mol. Biol. Rep. 2019, 46, 3713–3730. [Google Scholar] [CrossRef]
- Mishra, A.; Gupta, S.; Gupta, P.; Dhawan, S.S.; Lal, R.K. In Silico Identification of miRNA and Targets from Chrysopogon zizanioides (L.) Roberty with Functional Validation from Leaf and Root Tissues. Appl. Biochem. Biotechnol. 2020, 192, 1076–1092. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Avramenko, T.V. New opportunities for the regulation of secondary metabolism in plants: Focus on microRNAs. Biotechnol. Lett. 2015, 37, 1719–1727. [Google Scholar] [CrossRef]
- Paul, S.C.; Sharma, A.; Mehta, R.; Paul, S. In silico Characterization of microRNAs and Their Target Transcripts from Cranberry (Vaccinium macrocarpon). Cytol. Genet. 2020, 54, 82–90. [Google Scholar] [CrossRef]
- Paul, S.; de la Fuente-Jiménez, J.L.; Manriquez, C.G.; Sharma, A. Identification, characterization and expression analysis of passion fruit (Passiflora edulis) microRNAs. 3 Biotech 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Kundu, A.; Pal, A. Identification and expression profiling of Vigna mungo microRNAs from leaf small RNA transcriptome by deep sequencing. J. Integr. Plant Biol. 2014, 56, 15–23. [Google Scholar] [CrossRef]
- Zou, H.; Guo, X.; Yang, R.; Wang, S.; Li, L.; Niu, J.; Wang, D. MiR408- SmLAC3 Module Participates in Salvianolic Acid B Synthesis in Salvia miltiorrhiza. Int. J. Mol. Sci. 2021, 22, 7541. [Google Scholar] [CrossRef]
- Saifi, M.; Yogindran, S.; Nasrullah, N.; Nissar, U.; Gul, I.; Abdin, M.Z. Co-expression of anti-miR319g and miRStv-11 lead to enhanced steviol glycosides content in Stevia rebaudiana. BMC Plant Biol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bogdanović, M.; Cankar, K.; Dragićević, M.; Bouwmeester, H.; Beekwilder, J.; Simonović, A.; Todorović, S. Silencing of germacrene A synthase genes reduces guaianolide oxalate content in Cichorium intybus L. GM Crop. Food 2019, 11, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Kolanowska, M.; Kubiak, A.; Jażdżewski, K.; Wójcicka, A. MicroRNA analysis using next-generation sequencing. Methods Mol. Biol. 2018, 1823, 87–101. [Google Scholar] [CrossRef]
- Sharma, A.; Bejerano, P.I.A.; Maldonado, I.C.; de Donato Capote, M.; Madariaga-Navarrete, A.; Paul, S. Genome-wide computational prediction and experimental validation of quinoa (Chenopodium quinoa) microRNAs. Can. J. Plant Sci. 2019, 99, 666–675. [Google Scholar] [CrossRef]
- Kundu, A.; Paul, S.; Dey, A.; Pal, A. High throughput sequencing reveals modulation of microRNAs in Vigna mungo upon Mungbean Yellow Mosaic India Virus inoculation highlighting stress regulation. Plant Sci. 2017, 257, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.H.; Pan, X.P.; Cox, S.B.; Cobb, G.P.; Anderson, T.A. Evidence that miRNAs are different from other RNAs. Cell. Mol. Life Sci. 2006, 63, 246–254. [Google Scholar] [CrossRef]
- Liu, Y.C.; Chen, W.L.; Kung, W.H.; Huang, H. Da Plant miRNAs found in human circulating system provide evidences of cross kingdom RNAi. BMC Genomics 2017, 18, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lukasik, A.; Zielenkiewicz, P. Plant MicroRNAs-novel players in natural medicine? Int. J. Mol. Sci. 2017, 18, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Zhou, B.; Ross, S.A.; Zempleni, J. Nutrition, microRNAs, and Human Health. Adv. Nutr. 2017, 8, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Sabzehzari, M.; Naghavi, M.R. Phyto-miRNAs-based regulation of metabolites biosynthesis in medicinal plants. Gene 2019, 682, 13–24. [Google Scholar] [CrossRef]
- Yue, J.; Lu, X.; Zhang, H.; Ge, J.; Gao, X.; Liu, Y. Identification of conserved and novel microRNAs in blueberry. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kajal, M.; Singh, K. Small RNA profiling for identification of miRNAs involved in regulation of saponins biosynthesis in Chlorophytum borivilianum. BMC Plant Biol. 2017, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, M.P.; Pallaiyan, B.B.; Selvaraj, N. Chemical composition, antibacterial and antioxidant profile of essential oil from Murraya koenigii (L.) leaves. Avicenna J. Phytomed. 2014, 4, 200–214. [Google Scholar] [CrossRef]
- Sukkaew, S.; Pripdeevech, P.; Thongpoon, C.; Machan, T.; Wongchuphan, R. Volatile constituents of Murraya koenigii fresh leaves using headspace solid phase microextraction-gas chromatography-mass spectrometry. Nat. Prod. Commun. 2014, 9, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, R.S.; Chauhan, A.; Padalia, R.C.; Jat, S.K.; Thul, S.; Sundaresan, V. Phytochemical diversity of Murraya koenigii (L.) Spreng. from Western Himalaya. Chem. Biodivers. 2013, 10, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Meena, S.; Rajeev Kumar, S.; Dwivedi, V.; Kumar Singh, A.; Chanotiya, C.S.; Akhtar, M.Q.; Kumar, K.; Kumar Shasany, A.; Nagegowda, D.A. Transcriptomic insight into terpenoid and carbazole alkaloid biosynthesis, and functional characterization of two terpene synthases in curry tree (Murraya koenigii). Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakash, P.; Rajakani, R.; Gupta, V. Transcriptome-wide identification of Rauvolfia serpentina microRNAs and prediction of their potential targets. Comput. Biol. Chem. 2016, 61, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Srivastava, S.; Shasany, A.K.; Sharma, A. Identification of miRNAs and their targets involved in the secondary metabolic pathways of Mentha spp. Comput. Biol. Chem. 2016, 64, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.X.; Wang, L.J.; Zhao, B.; Shan, C.M.; Zhang, Y.H.; Chen, D.F.; Chen, X.Y. Progressive regulation of sesquiterpene biosynthesis in Arabidopsis and patchouli (Pogostemon cablin) by the MIR156-targeted SPL transcription factors. Mol. Plant 2015, 8, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Ashokkumar, K.; Selvaraj, K. Reverse phase-high performance liquid chromatography-diode array detector (RP-HPLC-DAD) analysis of flavonoids profile from curry leaf (Murraya koenigii. L). J. Med. Plants Res. 2013, 7, 3393–3399. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Karimi, E.; Rahmat, A. Optimization of ultrasound-assisted extraction of flavonoid compounds and their pharmaceutical activity from curry leaf (Murraya koenigii L.) using response surface methodology. BMC Complement. Altern. Med. 2014, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Zhang, X.; Luo, Z.; Zhang, Q.; Liu, J. Identification and characterization of microRNAs from Chinese pollination constant non-astringent persimmon using high-throughput sequencing. BMC Plant Biol. 2015, 15, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Tiwari, M.; Pandey, A.; Bhatia, C.; Sharma, A.; Trivedi, P.K. MicroRNA858 is a potential regulator of phenylpropanoid pathway and plant development. Plant Physiol. 2016, 171, 944–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Tang, Y.; Guo, Q.; Chen, J. Identification and characterization of microRNAs in phloem and xylem from ramie (Boehmeria nivea). Mol. Biol. Rep. 2020, 47, 1013–1020. [Google Scholar] [CrossRef]
- Li, H.; Dong, Y.; Yin, H.; Wang, N.; Yang, J.; Liu, X.; Wang, Y.; Wu, J.; Li, X. Characterization of the stress associated microRNAs in Glycine max by deep sequencing. BMC Plant Biol. 2011, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Anjali, N.; Nadiya, F.; Thomas, J.; Sabu, K.K. Identification and characterization of drought responsive microRNAs and their target genes in cardamom (Elettaria cardamomum Maton). Plant Growth Regul. 2019, 87, 201–216. [Google Scholar] [CrossRef]
- Shen, E.M.; Singh, S.K.; Ghosh, J.S.; Patra, B.; Paul, P.; Yuan, L.; Pattanaik, S. The miRNAome of Catharanthus roseus: Identification, expression analysis, and potential roles of microRNAs in regulation of terpenoid indole alkaloid biosynthesis. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Chen, T.; He, J.J.; Wu, J.H.; Luo, J.Y.; Ye, R.S.; Xie, M.Y.; Zhang, H.J.; Zeng, B.; Liu, J.; et al. Plant MIR167e-5p Inhibits Enterocyte Proliferation by Targeting β-Catenin. Cells 2019, 8, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, W.; Adolf, J.; Melzig, M.F. Identification of Viscum album L. miRNAs and prediction of their medicinal values. PLoS ONE 2017, 12, e0187776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qibin, L.; Jiang, W. MIREAP: microRNA Discovery by Deep Sequencing. 2008. Available online: https://sourceforge.net/projects/mireap/ (accessed on 10 June 2021).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Total Reads | Unique Reads |
|---|---|---|
| Total reads | 8186145 | 3726171 |
| Trimmed reads | 2052756 | 343505 |
| % reads aligned to ncRNA | 46.44% | |
| Reads aligned to ncRNA (rRNA, snoRNA, snRNA, tRNA) | 1,268,546 | 159,516 |
| Reads aligned to miRBase | 15,388 | 556 |
| Known miRNA | 142 | |
| Reads used for novel miRNA | 41,879 | 23,286 |
| Novel miRNA | 7 | |
| Putative miRNA | 726,943 | 79,115 |
| miRNA Family | Name | Sequence (5’-3’) | Length (nt) | Reference miRNA | No. of Mismatches | Read Counts | E Value |
|---|---|---|---|---|---|---|---|
| MIR156 | mko-miR156 | CTGACAGAAGAGAGTGAGCAC | 21 | ama-miR156 | 0 | 17 | 0.0000002 |
| mko-miR156 | TTGACGGAAGATAGAGAGCAC | 22 | bgy-miR156 | 1 | 6 | 0.00003 | |
| mko-miR156a | TGACAGAAGAGAGTGAACACA | 21 | bna-miR156a | 1 | 2 | 0.00004 | |
| mko-miR156a-3p | GCTCACTGCTCTTTCTGTCAG | 22 | ath-miR156a-3p | 0 | 7 | 0.0000001 | |
| mko-miR156d-3p | GCTCTCTATGCTTCTGTCATCA | 22 | stu-miR156d-3p | 0 | 1 | 0.00000004 | |
| mko-miR156e-5p | TGATAGAAGAGAGTGAGCA | 20 | sly-miR156e-5p | 0 | 1 | 0.000002 | |
| mko-miR156f-3p | TGCTCACTGCTCTTTCTG | 23 | bra-miR156f-3p | 0 | 4 | 0.000009 | |
| mko-miR156q | TGACAGAAGAGAGTGAGCACT | 21 | gma-miR156q | 0 | 39 | 0.0000002 | |
| mko-miR157d-3p | GCTCTCTATTCTTCTGTCATC | 21 | aly-miR157d-3p | 1 | 3 | 0.00004 | |
| MIR159 | mko-miR159 | TTTGGGTTGAAGGGAGCTCTA | 21 | pde-miR159 | 1 | 2 | 0.00004 |
| mko-miR159 | TTTGGACTGAAGGGAGCTCTA | 21 | aqc-miR159 | 0 | 1 | 0.0000002 | |
| mko-miR159a | TTTGGATTGAAGGGAGCTCTA | 21 | ath-miR159a | 0 | 1296 | 0.0000003 | |
| mko-miR159a-5p | GAGCTCCTTGAAGTCCAA | 21 | gma-miR159a-5p | 0 | 21 | 0.000009 | |
| mko-miR159b | TTGCATATCTCTGGAGCTTC | 21 | hbr-miR159b | 1 | 1 | 0.0001 | |
| mko-miR159b-3p | TTTGGATTGAAGGGAGCTCTT | 21 | ath-miR159b-3p | 0 | 29 | 0.0000002 | |
| mko-miR159c | TTTGGATTGAAGGGAGCTC | 21 | ath-miR159c | 0 | 4 | 0.000002 | |
| mko-miR159c | ATTGGATTGAAGGGAGCTC | 21 | osa-miR159c | 0 | 1 | 0.000002 | |
| mko-miR159c-5p | TTGGATCGAAGGGAGCTC | 21 | zma-miR159c-5p | 0 | 3 | 0.000009 | |
| mko-miR159d | AGCTGCTGAGCTATGGATCCC | 21 | gma-miR159d | 1 | 4 | 0.00009 | |
| mko-miR159e | TTTGGATTGAAAGGAGCTCT | 21 | sof-miR159e | 0 | 1 | 0.0000006 | |
| mko-miR159f | TTGGATTGAACGGAGCTCTA | 21 | osa-miR159f | 1 | 1 | 0.0001 | |
| MIR160 | mko-miR160e-5p | TGCCTGGCTCCCTGTATGCCG | 21 | osa-miR160e-5p | 0 | 14 | 0.0000002 |
| mko-miR160g | GCCTGGCTCCCTGTATGCCA | 23 | mes-miR160g | 0 | 30 | 0.0000009 | |
| MIR162_1 | mko-miR162b-3p | CGATAAACCTCTGCATCCAG | 21 | ath-miR162b-3p | 0 | 38 | 0.0000006 |
| MIR162_2 | mko-miR162b | TCGATAAGCCTCTGCATCCAG | 21 | osa-miR162b | 0 | 2 | 0.0000002 |
| MIR164 | mko-miR164b | TGGAGAAGCAGGGCACGT | 20 | gma-miR164b | 0 | 68 | 0.000009 |
| mko-miR164b-5p | TGGAGAAGTAGGGCACGTGCA | 21 | ath-miR164b-5p | 1 | 2 | 0.00004 | |
| mko-miR164d | TGGAGAAGCAGGGCACATGCT | 21 | mtr-miR164d | 0 | 2 | 0.0000002 | |
| MIR166 | mko-miR165b | TCGGACCAGGCTTCATCCCC | 21 | ath-miR165b | 0 | 2 | 0.0000006 |
| mko-miR166a | TCGGACCAGGCTTCATTCCCC | 21 | pta-miR166a | 0 | 2258 | 0.0000003 | |
| mko-miR166b | TCGGACCAGGCTTCATTCCCT | 22 | crt-miR166b | 0 | 362 | 0.0000002 | |
| mko-miR166b | TCGGACCAGGCTTCATTCCT | 21 | mtr-miR166b | 0 | 5 | 0.0000006 | |
| mko-miR166b | TCGGANCAGGCTTCATTCCCG | 22 | csi-miR166b | 1 | 1 | 0.000009 | |
| mko-miR166c-5p | GGAATGTTGTCTGGCTCGAGG | 21 | gma-miR166c-5p | 0 | 144 | 0.0000003 | |
| mko-miR166d | TCGGGCCAGGCTTCATTCCCC | 21 | mtr-miR166d | 1 | 2 | 0.00004 | |
| mko-miR166e-3p | TCGAACCAGGCTTCATTCCCC | 21 | osa-miR166e-3p | 0 | 2 | 0.0000002 | |
| mko-miR166h-5p | GGAATGTTGTTTGGCTCGAGG | 21 | gma-miR166h-5p | 0 | 6 | 0.0000001 | |
| mko-miR166i | TCGGACCAGGCTTCATTCT | 20 | cme-miR166i | 0 | 2 | 0.000002 | |
| mko-miR166i-3p | TCGGATCAGGCTTCATTCC | 21 | osa-miR166i-3p | 0 | 1 | 0.000002 | |
| mko-miR166k | TCTCGGACCAGGCTTCGTTCC | 21 | gma-miR166k | 1 | 2 | 0.00004 | |
| mko-miR166k-3p | CGGACCAGGCTTCAATCCC | 21 | osa-miR166k-3p | 0 | 2 | 0.000002 | |
| mko-miR166m | CGGACTAGGCTTCATTCCCC | 20 | gma-miR166m | 1 | 1 | 0.0001 | |
| mko-miR166p | TCGGACCAGGCTCCATTCC | 21 | ptc-miR166p | 0 | 2 | 0.000002 | |
| mko-miR166u | TCTCGGACCAGGCTTCATT | 20 | gma-miR166u | 0 | 3 | 0.000002 | |
| MIR167_1 | mko-miR167a | AGATCATCTGGCAGTTTCACC | 21 | mdm-miR167a | 0 | 78 | 0.0000002 |
| mko-miR167b | TGAAGCTGACAGCATGATCT | 21 | tae-miR167b | 0 | 2 | 0.0000006 | |
| mko-miR167b | TGAAGCTGCCAGCATGATCTA | 22 | bna-miR167b | 0 | 1 | 0.0000001 | |
| mko-miR167c-5p | TGAAGCTGCCAGCATGATCTGC | 22 | tae-miR167c-5p | 0 | 1047 | 0.00000004 | |
| mko-miR167d | TGAAGCTGCCAGCATGATCTGA | 22 | cpa-miR167d | 0 | 19 | 0.00000004 | |
| mko-miR167d | TGAAGCAGCCAGCATGATCTGG | 22 | ath-miR167d | 1 | 3 | 0.000009 | |
| mko-miR167d | TGAAGCTGCCATCATGATCT | 20 | bna-miR167d | 1 | 2 | 0.0001 | |
| mko-miR167f-3p | ATCATGTGGCAGTTTCACC | 21 | ptc-miR167f-3p | 0 | 3 | 0.000002 | |
| mko-miR167h-5p | TGAAGCTGCCAACATGATCTG | 21 | ptc-miR167h-5p | 0 | 1 | 0.0000001 | |
| mko-miR167i | TGAAGCTGCCAGCAAGATCTTA | 22 | mdm-miR167i | 1 | 1 | 0.000009 | |
| MIR168 | mko-miR168 | TCCCGCCTTGCATCAACTG | 24 | aau-miR168 | 0 | 2 | 0.000002 |
| mko-miR168a-3p | CCCGCCTTGCATCAACTGAAT | 21 | ath-miR168a-3p | 0 | 9 | 0.0000001 | |
| mko-miR168b | TCGCTTGGTGCAGGTCGGG | 19 | gma-miR168b | 0 | 9 | 0.000002 | |
| mko-miR168b-5p | TCGCCTGGTGCAGGTCGGGAA | 21 | ath-miR168b-5p | 1 | 2 | 0.00004 | |
| MIR169_1 | mko-miR169h | TAGCCAAGGATGACTTGCCTG | 21 | ath-miR169h | 0 | 1 | 0.0000002 |
| MIR171_1 | mko-miR171a | TTGAGCCGCGTCAATATCTCC | 21 | mdm-miR171a | 0 | 7 | 0.0000002 |
| MIR172 | mko-miR172b | GTGTAGCATCATCAAGAT | 21 | vvi-miR172b | 0 | 4 | 0.000009 |
| mko-miR172b-5p | GCAGCACCATCAAGATTCACA | 21 | aly-miR172b-5p | 0 | 18 | 0.0000002 | |
| mko-miR172c-5p | GTAGCATCATCAAGATTCACA | 21 | mtr-miR172c-5p | 0 | 7 | 0.0000002 | |
| MIR390 | mko-miR390a-3p | CGCTATCCATCCTGAGTTTCA | 21 | ath-miR390a-3p | 0 | 5 | 0.0000002 |
| mko-miR390b | AAGCTCAGGAGGGATAGCGCC | 21 | ppt-miR390b | 0 | 5 | 0.0000002 | |
| MIR393 | mko-miR393 | CCAAAGGGATCGCATTGATCT | 22 | ghr-miR393 | 0 | 3 | 0.0000001 |
| MIR396 | mko-miR396 | TTCCACAGCTTTCTTGAACTT | 21 | pta-miR396 | 0 | 35 | 0.0000002 |
| mko-miR396-3p | GCTCAAGAAAGCTGTGGGA | 21 | ama-miR396-3p | 0 | 2 | 0.000002 | |
| mko-miR396a | CACAGCTTTCTTGAACTT | 21 | hbr-miR396a | 0 | 1 | 0.000009 | |
| mko-miR396a-3p | GTTCAATAAAGCTGTGGGAAG | 21 | ath-miR396a-3p | 0 | 261 | 0.0000003 | |
| mko-miR396a-3p | TTCAATAAAGCTGAGGGAAG | 20 | gma-miR396a-3p | 1 | 1 | 0.0001 | |
| mko-miR396a-5p | TTCCACAGCTTTCTTGAACTG | 21 | ath-miR396a-5p | 0 | 1121 | 0.0000002 | |
| mko-miR396b-3p | GTTCAATAAAGCTGTGGGA | 20 | osa-miR396b-3p | 0 | 4 | 0.000003 | |
| mko-miR396c | TTCAAGAAATCTGTGGGAAG | 20 | csi-miR396c | 0 | 22 | 0.0000006 | |
| mko-miR396g-3p | CTCAAGAATGCCGTGGGAAA | 21 | ptc-miR396g-3p | 1 | 3 | 0.0001 | |
| mko-miR396g-3p | GTTCAAGAAAGCTGTGGAAG | 21 | zma-miR396g-3p | 0 | 2 | 0.0000007 | |
| mko-miR396g-5p | TTCCACGGCTTTCTTGAACTT | 21 | ptc-miR396g-5p | 0 | 144 | 0.0000001 | |
| mko-miR396j | TTCCACAGCTATCTTGAA | 21 | gma-miR396j | 0 | 2 | 0.000009 | |
| MIR399 | mko-miR399e | CGCCAAAGGAGAGTTGCCCT | 21 | ptc-miR399e | 0 | 1 | 0.0000006 |
| MIR403 | mko-miR403-3p | TTAGATTCACGCACAAACTCG | 21 | ath-miR403-3p | 0 | 36 | 0.0000002 |
| MIR408 | mko-miR408-5p | GGGGAACAGGCAGAGCATGG | 21 | ptc-miR408-5p | 0 | 2 | 0.0000006 |
| MIR408_2 | mko-miR408 | ATGCACTGCCTCTTCCCTGGC | 21 | pta-miR408 | 0 | 1 | 0.0000002 |
| MIR477 | mko-miR477a | ACTCTCCCTCAAGGGCTTCTG | 21 | nta-miR477a | 0 | 6 | 0.0000002 |
| mko-miR477a | ATCTCCCTTAAAGGCTTCCAA | 21 | vvi-miR477a | 1 | 2 | 0.00003 | |
| MIR482 | mko-miR472 | TTTTCCCACACCTCCCATCCC | 21 | csi-miR472 | 0 | 36 | 0.0000001 |
| mko-miR482 | TCTTCCCTACTCCACCCAT | 22 | mes-miR482 | 0 | 99 | 0.000002 | |
| mko-miR482a-3p | TCTTCCCTAAGCCTCCCATTCC | 22 | csi-miR482a-3p | 1 | 2 | 0.000009 | |
| mko-miR482b | TCTTGCCCAACCCTCCCATTCC | 22 | csi-miR482b | 1 | 192 | 0.000009 | |
| mko-miR482b | TTGCCAACTCCACCCATGCC | 22 | ghr-miR482b | 1 | 1 | 0.0001 | |
| mko-miR482c | TTCCCTAGTCCCCCTATTCCTA | 22 | csi-miR482c | 0 | 138 | 0.00000004 | |
| mko-miR482d-3p | TCTTCCCTACACCACCCAT | 22 | gma-miR482d-3p | 1 | 1 | 0.0005 | |
| MIR530 | mko-miR530-3p | AGGTGCAGAGGCAGATGCAAC | 21 | osa-miR530-3p | 0 | 1 | 0.0000002 |
| MIR535 | mko-miR535 | TGACAATGAGAGAGAGCAC | 21 | csi-miR535 | 0 | 422 | 0.000005 |
| mko-miR535b | TGACAAAGAGAGAGAGCACGC | 21 | mdm-miR535b | 1 | 3 | 0.00003 | |
| mko-miR535d | TGACGATGAGAGAGAGCACGC | 21 | mdm-miR535d | 1 | 1 | 0.00004 | |
| mko-miR535d | TGACAACGAGAGAGAGCACGC | 21 | ppt-miR535d | 0 | 1 | 0.0000001 | |
| MIR827 | mko-miR827 | TTTGCTGATTGTCATCTAA | 21 | osa-miR827 | 1 | 1 | 0.0006 |
| mko-miR827-5p | TTTGTTGATTGTCATCTAA | 22 | bdi-miR827-5p | 1 | 16 | 0.0006 | |
| MIR827_2 | mko-miR827b | TTAGATGACCATCAACAAACA | 21 | ghr-miR827b | 0 | 21 | 0.0000002 |
| MIR845_2 | mko-miR845e | TGGCTCTGATACCAATTGATG | 21 | vvi-miR845e | 0 | 1 | 0.0000002 |
| MIR845_3 | mko-miR845b | GCTCTAATACCAATTGATA | 21 | vvi-miR845b | 1 | 3 | 0.0006 |
| MIR858 | mko-miR858 | CTCGTTGTCTGTTCGACCTTG | 21 | ppe-miR858 | 0 | 82 | 0.0000002 |
| MIR1446 | mko-miR1446 | AACTCTCTCCCTCATAGGCT | 21 | gra-miR1446 | 1 | 5 | 0.0001 |
| MIR1507 | mko-miR1507-3p | CCTCGTTCCAAACATCATCT | 22 | mtr-miR1507-3p | 1 | 3 | 0.0001 |
| MIR2673 | mko-miR2673b | GAAGAGGAAGAGGAAGAGG | 22 | mtr-miR2673b | 0 | 1 | 0.000005 |
| MIR3630 | mko-miR3630-3p | TGGGAATCTCTCTGATGCA | 22 | vvi-miR3630-3p | 0 | 2 | 0.000004 |
| MIR8654 | mko-miR8654c | AGGATACTGCTTTGATGGA | 24 | gra-miR8654c | 1 | 2 | 0.0007 |
| MIR9560 | mko-miR9560a-5p | CAGGAGGTGGAACAAATATGA | 24 | bra-miR9560a-5p | 1 | 19 | 0.00004 |
| NA | mko-miR156i | GACAGAAAAGAGAGAGCAG | 20 | ath-miR156i | 1 | 1 | 0.0005 |
| mko-miR477b | CTCTCCCTCAAGGGCTTCT | 21 | nta-miR477b | 0 | 2 | 0.000002 | |
| mko-miR477h | ACTCTCCCTCAAGGGCTTCA | 21 | mes-miR477h | 0 | 1 | 0.0000006 | |
| mko-miR845 | GCTCTGATACCAATTGTTG | 21 | bdi-miR845 | 0 | 4 | 0.000003 | |
| mko-miR894 | CGTTTCACGTCGGGTTCACC | 20 | ppt-miR894 | 0 | 2 | 0.0000006 | |
| mko-miR1446 | TTCTAAACTCTCTCCCTCAT | 20 | mes-miR1446 | 1 | 3 | 0.0001 | |
| mko-miR1515 | ATTTTTGCGTGCAATGATCC | 22 | csi-miR1515 | 0 | 2 | 0.0000006 | |
| mko-miR2916 | TGGGGGCTCGAAGACGATCA | 23 | peu-miR2916 | 1 | 13 | 0.0003 | |
| mko-miR3711 | GCCCTCCTTCTAGCGCCA | 20 | pab-miR3711 | 0 | 23 | 0.00002 | |
| mko-miR3948 | TGGAGTGGGAGTGAGAGTA | 24 | csi-miR3948 | 1 | 1 | 0.0006 | |
| mko-miR3950 | AGAAATCATGTTGCAGAAA | 21 | csi-miR3950 | 1 | 1 | 0.0006 | |
| mko-miR3952 | TGAAGGGCCTTTCTAGAGCAC | 21 | csi-miR3952 | 0 | 8 | 0.0000002 | |
| mko-miR3954 | TGGACAGAGAAATCACGGTCA | 21 | csi-miR3954 | 0 | 19 | 0.0000001 | |
| mko-miR4995 | AGGCAGTGGCTTGGTTAAGGG | 21 | gma-miR4995 | 0 | 1 | 0.0000003 | |
| mko-miR5021 | GAGAAGAAGAAGAAGAAAA | 20 | ath-miR5021 | 0 | 1 | 0.000002 | |
| mko-miR5072 | TTCCCCAGTGGAGTCGCCA | 22 | osa-miR5072 | 1 | 2 | 0.0006 | |
| mko-miR5082 | ATGATGGCCTCGCGGGTTCA | 24 | osa-miR5082 | 1 | 47 | 0.0002 | |
| mko-miR5083 | AGACTACAATTATCTGATCA | 20 | osa-miR5083 | 0 | 27 | 0.000001 | |
| mko-miR5368 | GGACAGTCTCAGGTAGACA | 19 | gma-miR5368 | 0 | 592 | 0.000005 | |
| mko-miR5523 | CTAGTAAATACGTTCCTCCTCA | 22 | osa-miR5523 | 1 | 22 | 0.00002 | |
| mko-miR5532 | ATGGAATATATGACAAGGGTGG | 22 | osa-miR5532 | 1 | 2 | 0.00002 | |
| mko-miR5538 | GCAGCAAGTGATTGAGTTCAGT | 22 | osa-miR5538 | 0 | 2 | 0.00000009 | |
| mko-miR5658 | ATGATGAGGATGATGATGAA | 21 | ath-miR5658 | 1 | 2 | 0.0003 | |
| mko-miR5817 | GAATTTGAAAAAAAAAGGT | 24 | osa-miR5817 | 1 | 2 | 0.0009 | |
| mko-miR6173 | AGCCGTAAACGATGGATACT | 20 | hbr-miR6173 | 0 | 3 | 0.000001 | |
| mko-miR6300 | GTCGTTGTAGTATAGTGG | 18 | gma-miR6300 | 0 | 803 | 0.00002 | |
| mko-miR6478 | CCGACCTTAGCTCAGTTGGT | 21 | ptc-miR6478 | 0 | 9 | 0.000001 | |
| mko-miR6483 | TATTGTAGAAATTTTCGGGATC | 22 | hbr-miR6483 | 1 | 19 | 0.00002 | |
| mko-miR6485 | TAGGATGTAGAAGATCATAA | 20 | hbr-miR6485 | 1 | 4 | 0.0003 | |
| mko-miR7767-5p | CCCCAAGATGAGAGCTCTCC | 21 | bdi-miR7767-5p | 1 | 1 | 0.0003 | |
| mko-miR8051-5p | TGAATCTTTATACCATACTA | 20 | stu-miR8051-5p | 1 | 14 | 0.0003 | |
| mko-miR8175 | GATCCCCGGCAACGGCGCCA | 20 | ath-miR8175 | 0 | 661 | 0.000001 | |
| mko-miR8610.1 | TTTTCTGAACAAATCGAAGAA | 24 | atr-miR8610.1 | 1 | 44 | 0.00009 | |
| mko-miR9774 | GAAATACCCAATATCTTG | 22 | tae-miR9774 | 0 | 1 | 0.000009 |
| Name | Sequence (5′-3′) | Length | Read Count | Strand | MFEI of Precursor |
|---|---|---|---|---|---|
| mko-miRN1-3p | UUAGGGUUUCAGUGAUCGAAAAC | 23 | 14 | − | 0.85 |
| mko-miRN2-3p | GUGAGCCAAGCAAGUAGUGUCGC | 23 | 5 | + | 0.70 |
| mko-miRN3-3p | UUGUCUCACUGCCUGUUGCACU | 22 | 7 | + | 0.92 |
| mko-miRN4-5p | UGCAGGUGAGAUGAUACCGUCA | 22 | 15 | − | 0.72 |
| mko-miRN5-3p | ACCGUGUUUCUCUGCCCAAUCAG | 23 | 8 | − | 0.98 |
| mko-miRN6-5p | CUGGGGAGUUGCACCCGGAGUA | 22 | 6 | − | 0.79 |
| mko-miRN7-3p | UUGUUUUGGGUGAAACGGGUGUU | 23 | 93 | + | 0.97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-García, C.; Ahmed, S.S.S.J.; Ramalingam, S.; Selvaraj, D.; Srivastava, A.; Paul, S.; Sharma, A. Identification of microRNAs from Medicinal Plant Murraya koenigii by High-Throughput Sequencing and Their Functional Implications in Secondary Metabolite Biosynthesis. Plants 2022, 11, 46. https://doi.org/10.3390/plants11010046
Gutiérrez-García C, Ahmed SSSJ, Ramalingam S, Selvaraj D, Srivastava A, Paul S, Sharma A. Identification of microRNAs from Medicinal Plant Murraya koenigii by High-Throughput Sequencing and Their Functional Implications in Secondary Metabolite Biosynthesis. Plants. 2022; 11(1):46. https://doi.org/10.3390/plants11010046
Chicago/Turabian StyleGutiérrez-García, Claudia, Shiek S. S. J. Ahmed, Sathishkumar Ramalingam, Dhivya Selvaraj, Aashish Srivastava, Sujay Paul, and Ashutosh Sharma. 2022. "Identification of microRNAs from Medicinal Plant Murraya koenigii by High-Throughput Sequencing and Their Functional Implications in Secondary Metabolite Biosynthesis" Plants 11, no. 1: 46. https://doi.org/10.3390/plants11010046