
First Estimates of Age and Growth of the Lusitanian Cownose Ray (Rhinoptera marginata) from the Mediterranean Sea


Abstract
:1. Introduction
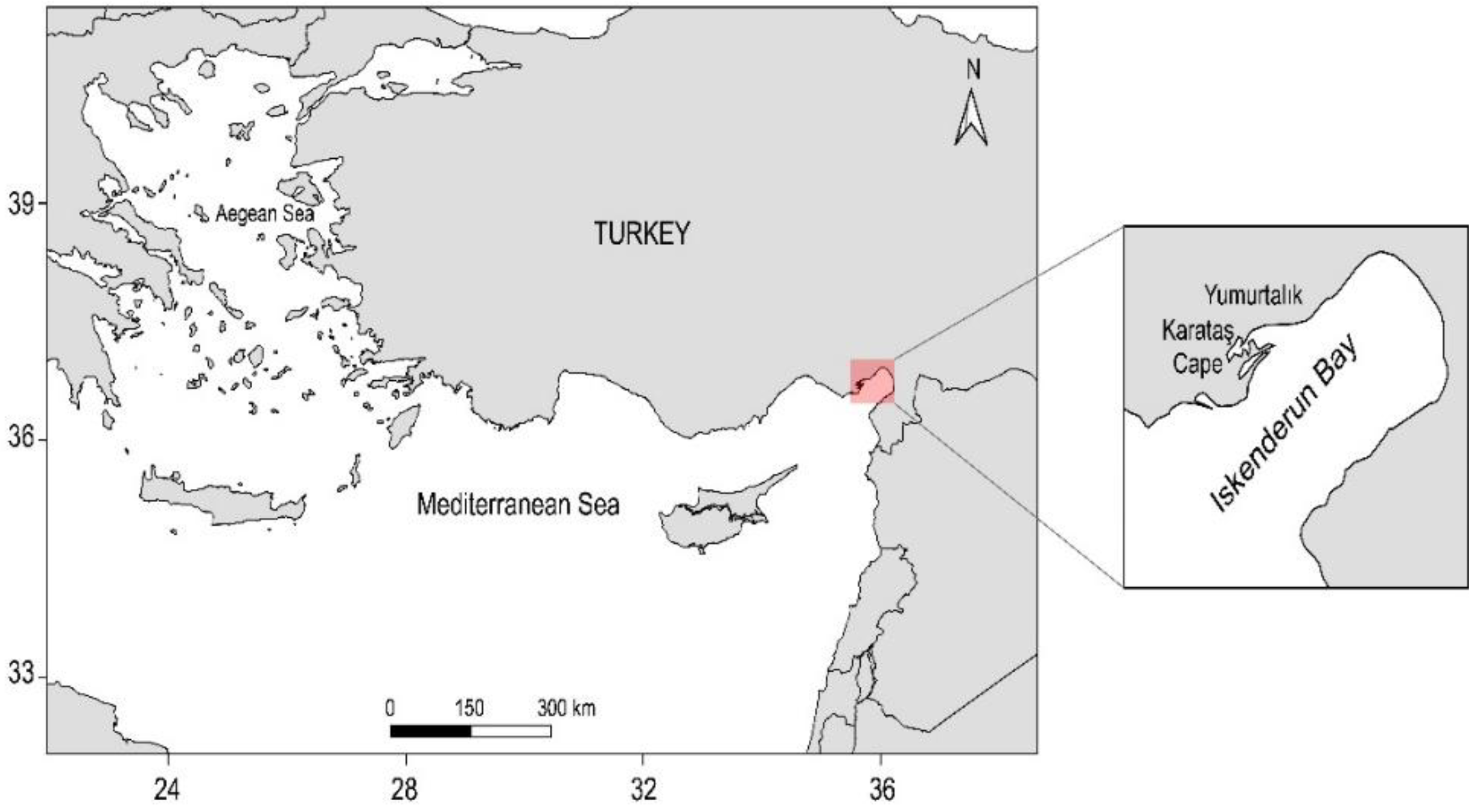
2. Materials and Methods
3. Results
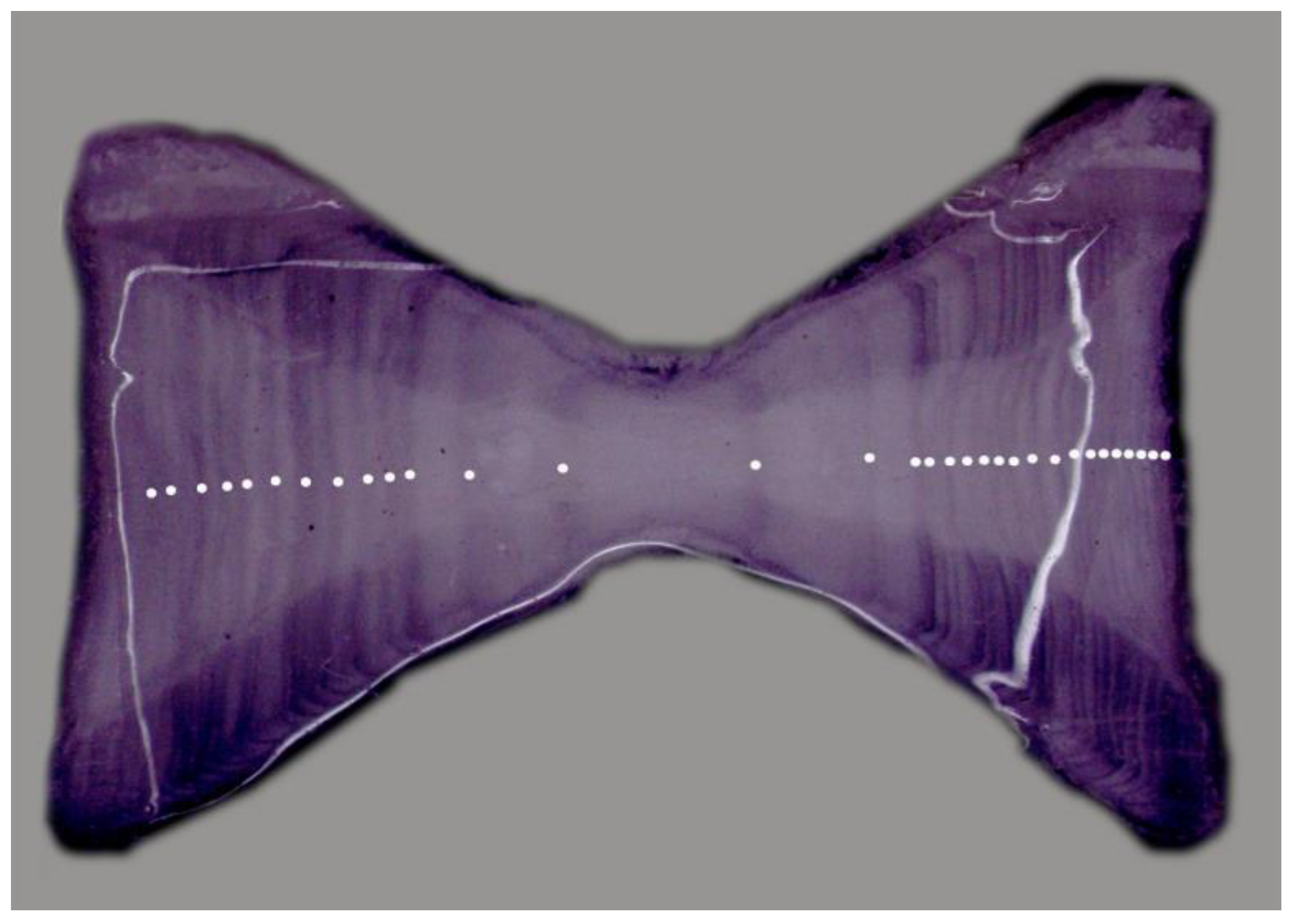
3.1. Size Composition and Vertebral Analysis
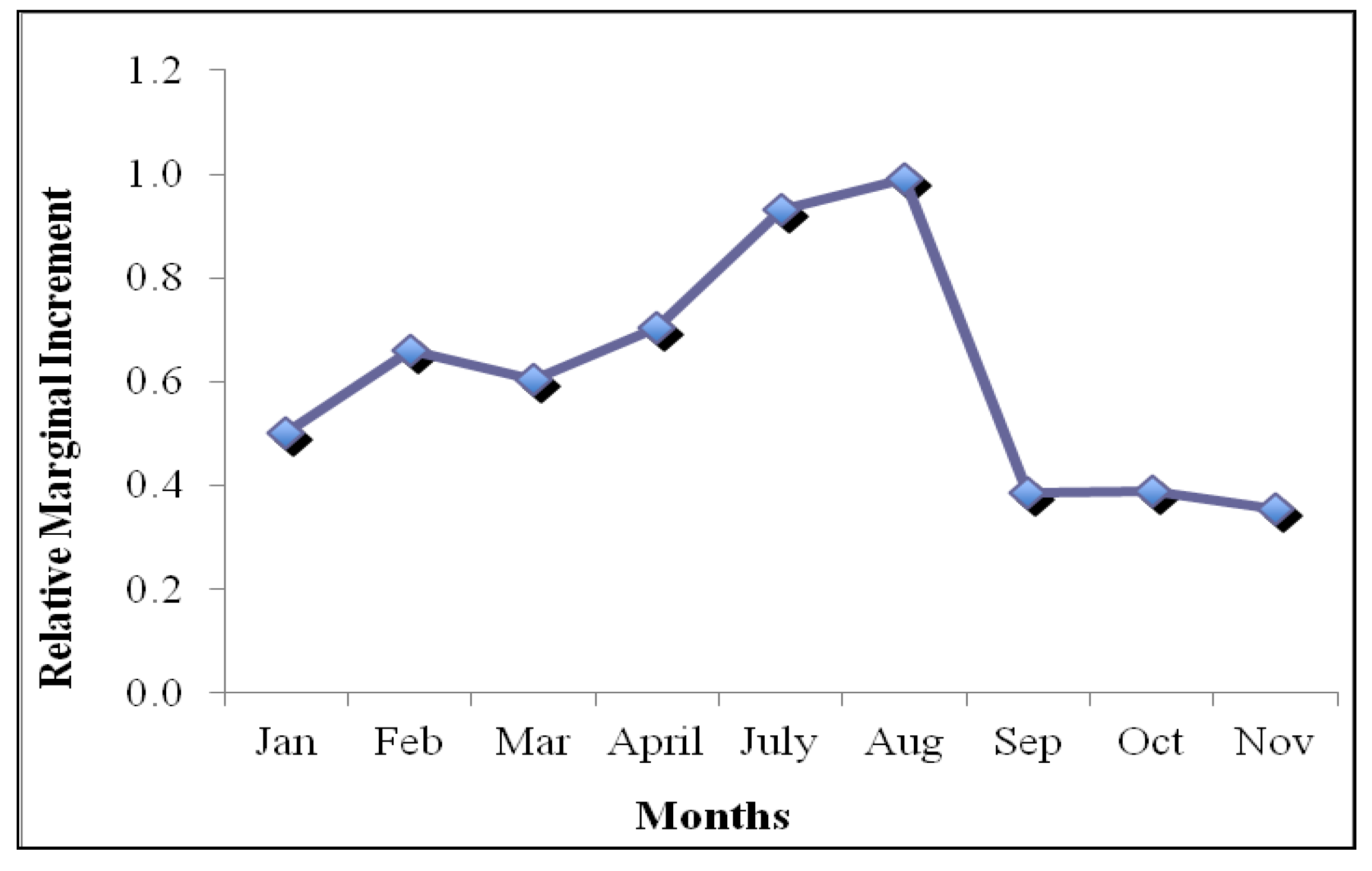
3.2. Marginal Increment Analysis
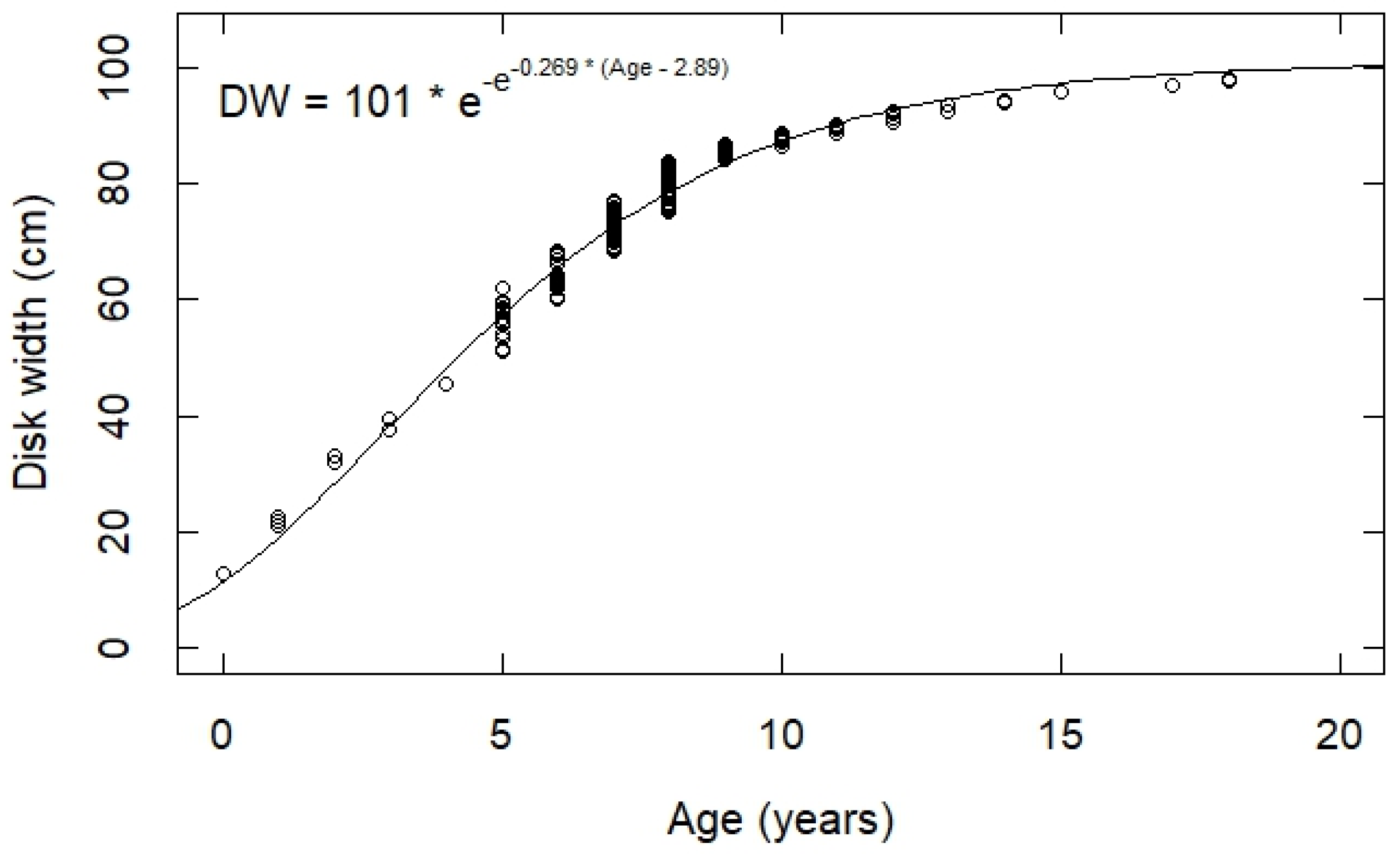
3.3. Back Calculation and Growth Estimates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Golani, D.; Öztürk, B.; Başusta, N. Fishes of the Eastern Mediterranean, 1st ed.; Turkish Marine Research Foundation: Istanbul, Turkey, 2006; p. 259. [Google Scholar]
- Başusta, A.; Özer, E.I.; Sulikowski, J.A.; Başusta, N. First record of a gravid female and neonate of the Lusitanian cownose ray, Rhinoptera marginata, from the eastern Mediterranean Sea. J. Appl. Ichthyol. 2012, 28, 643–644. [Google Scholar] [CrossRef]
- Campredon, P.; Cuq, F. Artisanal fishing and coastal conservation in West Africa. J. Coast Conserv. 2001, 7, 91–100. [Google Scholar] [CrossRef]
- Yaglioglu, D.; Gurlek, M.; Turan, C. On the occurrence of the giant devil ray Mobula mobular (Bonnaterre, 1788) from the Mediterranean coast of Turkey-by-catch documentation. J. Appl. Ichthyol. 2013, 29, 935–936. [Google Scholar] [CrossRef]
- Notarbartolo di Sciara, G.; Serena, F.; Ducrocq, M.; Seret, B. Rhinoptera marginata (Lusitanian Cownose Ray). In ‘The IUCN Red List of Threatened Species’. (International Union for Conservation of Nature and Natural Resources). 2009. Available online: http://www.iucnredlist.org/details/161463/0 (accessed on 23 November 2017).
- Başusta, N.; Demirhan, S.A.; Çiçek, E.; Başusta, A.; Kuleli, T. Age and growth of the common guitarfish, Rhinobatos rhinobatos (Linnaeus, 1758), in Iskenderun Bay (northeastern Mediterranean, Turkey). J. Mar. Biol. Assoc. UK 2008, 88, 837–842. [Google Scholar] [CrossRef]
- Çek, Ş.; Başusta, N.; Demirhan, S.A.; Karalar, M. Biological observations on the common guitarfish (Rhinobatos rhinobatos Linnaeus 1758) from Iskenderun Bay. Anim. Biol. 2009, 59, 211–230. [Google Scholar] [CrossRef]
- Ceyhan, T.; Hepkafadar, O.; Tosunoglu, Z. Catch and size selectivity of small-scale fishing gear for the smooth-hound shark Mustelus mustelus (Linnaeus, 1758) (Chondrichthyes: Triakidae) from the Aegean Turkish coast. Mediterr. Mar. Sci. 2010, 11, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Başusta, N.; Erdem, U. A study on the pelagic and demersal fishes in Iskenderun Bay. Turk. J. Zool. 2000, 24, 1–19, (In Turkish with English Abstract). [Google Scholar]
- Quignard, J.P.; Tomasini, J.A. Mediterranean fish biodiversity. Biol. Mar. Mediterr. 2000, 7, 1–66. [Google Scholar]
- Bilecenoglu, M.; Taskavak, E.; Mater, S.; Kaya, M. Checklist of the marine fishes of Turkey. Zootaxa 2002, 113, 1–194. [Google Scholar] [CrossRef]
- Başusta, A.; Basusta, N.; Sulikowski, J.A.; Driggers, W.B.; Demirhan, S.A.; Cicek, E. Length-weight relationships for nine species of batoids from the Iskenderun Bay, Turkey. J. Appl. Ichthyol. 2012, 28, 850–851. [Google Scholar] [CrossRef]
- Tiraşin, E.M.; Başusta, N. Near-term embryos and gravid females of Lusitanian cownose ray (Rhinoptera marginata) in Mersin Bay, eastern Mediterranean Sea. Mar. Freshw. Res. 2018, 69, 1365–1371. [Google Scholar] [CrossRef]
- Jennings, S.; Kaiser, M.J.; Reynolds, J.D. Marine Fisheries Ecology, 1st ed.; Blackwell Publishing: Malden, MA, USA, 2001; pp. 178–204. [Google Scholar]
- Cailliet, G.M.; Goldman, K.J. Age determination and validation in chondrichthyan fishes. In The Biology of Sharks and Their Relatives; Carrier, J., Musick, J.A., Heithaus, M., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 399–447. [Google Scholar]
- McFarlane, G.A.; King, R. Age and growth of big skate (Raja binoculata) and longnose skate (Raja rhina) in British Columbia waters. Fish. Res. 2006, 78, 169–178. [Google Scholar] [CrossRef]
- Bizzarro, J.J.; Smith, W.D.; M’arquez-Farias, J.F.; Hueter, R.E. Artisanal fisheries and reproductive biology of the golden cownose ray, Rhinoptera steindachneri Evermann and Jenkins, 1891, in the northern Mexican Pacific. Fish. Res. 2007, 84, 137–146. [Google Scholar] [CrossRef]
- Hoff, T.B.; Musick, J.A. Western North Atlantic shark-fishery management problems and informational requirements. In Elasmobranchs as Living Resources: Advances in the Biology, Ecology, Systematics, and the Status of Fisheries; NOAA Technical Report, NMFS 90; Pratt, H.L., Jr., Gruber, S.H., Taniuchi, T., Eds.; U.S. Department of Commerce: Washington, DC, USA, 1990; pp. 455–472. [Google Scholar]
- Sulikowski, J.A.; Morin, M.D.; Suk, S.H.; Howell, W.H. Age and growth estimates of the winter skate (Leucoraja ocellata) in the western gulf of Maine. Fish. Bull. 2003, 101, 405–413. [Google Scholar]
- Girgin, H.; Başusta, N. Testing staining techniques to determine age and growth of Dasyatis pastinaca (Linnaeus, 1758) captured in Iskenderun Bay, northeastern Mediterranean. J. Appl. Ichthyol. 2016, 32, 595–601. [Google Scholar] [CrossRef]
- Beamish, R.J.; Fournier, D.A. A method for comparing the precision of a set of age determinations. Can. J. Fish. Aquat. Sci. 1981, 38, 982–983. [Google Scholar] [CrossRef]
- Campana, S.E. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Cailliet, G.M. Elasmobranch age determination and verification: An updated review. In Elasmobranchs as Living Resources: Advances in the Biology, Ecology, Systematics, and the Status of Fisheries; NOAA Technical Report, NMFS 90; Pratt, H.L., Jr., Gruber, S.H., Taniuchi, T., Eds.; U.S. Department of Commerce: Washington, DC, USA, 1990; pp. 157–165. [Google Scholar]
- Sulikowski, J.A.; Irvine, S.B.; DeValerio, K.C.; Carlson, J.K. Age growth and maturity of the roundel skate, Raja texana, from the Gulf of Mexico, USA. Mar. Freshw. Res. 2009, 58, 41–53. [Google Scholar] [CrossRef]
- von Bertalanffy, L. A quantitative theory of organic growth (inquiries on growth laws. II). Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Richer, W.E. Computation and Interpretation of Biological Statistics of Fish Populations; Bullettin of the Fisheries Research Board Canada, 191; Department of the Environment Fisheries and Marine Service: Ottowa, ON, Canada, 1975; p. 382.
- Tirasin, E.M. Balık populasyonlarının büyüme parametrelerinin araştırılması. Turk. J. Zool. 1993, 17, 29–82, (In Turkish with English Abstract). [Google Scholar]
- Walker, P.; Cavanagh, R.D.; Ducrocq, M.; Fowler, S.L. Regional Overviews: Northeast Atlantic (including Mediterranean and Black Sea). In Sharks, Rays and Chimaeras: The Status of the Chondrichthyan Fishes; Fowler, S.L., Cavanagh, R.D., Camhi, M., Burgess, G.H., Cailliet, G.M., Fordham, S.V., Simpfendorfer, C.A., Musick, J.A., Eds.; IUCN SSC Shark Specialist Group, IUCN: Gland, Switzerland; Cambridge, UK, 2005; pp. 71–86. [Google Scholar]
- Cavanagh, R.D.; Gibson, C. Overview of the Conservation Status of Cartilaginous Fishes (Chondrichthyans) in the Mediterranean Sea; IUCN: Gland, Switzerland; Malaga, Spain, 2007; p. 42. [Google Scholar]
- Worm, B.; Davis, B.; Kettemer, L.; Ward-Paige, C.A.; Chapman, D.; Heithaus, M.R.; Kessel, S.T.; Gruber, S.H. Global catches, exploitation rates, and rebuilding options for sharks. Mar. Policy 2013, 40, 194–204. [Google Scholar] [CrossRef]
- Smith, J.W.; Merriner, J.V. Age and growth, movements and distribution of the cownose ray, Rhinoptera bonasus, in Chesapeake Bay. Estuaries 1987, 10, 153–164. [Google Scholar] [CrossRef]
- Neer, J.A.; Thompson, B.A. Life history of the cownose ray, Rhinoptera bonasus, in the northern Gulf of Mexico, with comments on geographic variability in life history traits. Environ. Biol. Fishes 2005, 73, 321–331. [Google Scholar] [CrossRef]
- Fisher, R.A.; Call, G.C.; Grubbs, R.D. Age, growth, and reproductive biology of cownose rays in Chesapeake Bay. Mar. Coast Fish. 2013, 5, 224–235. [Google Scholar] [CrossRef] [Green Version]
- Last, P.R.; De Carvalho, M.R.; Corrigan, S.; Naylor, G.J.P.; Seret, B.; Yang, L. The rays of the world project—An explanation of nomenclatural decisions. In Rays of the World, Supplementary Information; Last, P.R., Yearsley, G.K., Eds.; CSIRO Australian National Fish Collection: Clayton, Australia, 2016; pp. 1–10. [Google Scholar]
- Ebert, D.A.; Stehmann, M.F.W. Sharks, Batoids, and Chimaeras of the North Atlantic; FAO Species Catalogue for Fishery Purposes, 7; FAO: Rome, Italy, 2013; p. 523. [Google Scholar]
- Neer, J.A.; Cailliet, G.M. Aspects of the life history of the pacific electric ray, Torpedo californica, Ayres. Copeia 2001, 3, 842–847. [Google Scholar] [CrossRef]
- Mollet, H.F.; Ezcurra, J.M.; O’sullivan, J.B. Captive biology of the pelagic stingray, Dasyatis violacea (Bonaparte, 1832). Mar. Freshw. Res. 2002, 53, 531–541. [Google Scholar] [CrossRef]
- Martin, L.K.; Cailliet, G.M. Age and growth determination of the bat ray, Myliobatis california Gill, in central California. Copeia 1988, 3, 762–773. [Google Scholar] [CrossRef]
- Coutre, K.; Gedamke, T.; Rudders, D.B.; Driggers, W.B.; Koester, D.M.; Sulikowski, J.A. Indication of density-dependent changes in growth and maturity of the barndoor skate on Georges Bank. Mar. Coast Fish. 2013, 5, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Mejía-Falla, P.A.; Cortés, E.; Navia, A.F.; Zapata, F.A. Age and growth of the round stingray Urotrygon rogersi, a particularly fast-growing and short-lived elasmobranch. PLoS ONE 2014, 9, e96077. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Female DW (cm) | Female W (g) | Male DW (cm) | Male W (g) |
|---|---|---|---|---|
| 0 | - | - | 13.0 (1) | 172 |
| 1 | 21.4 ± 5 (2) | 270 ± 45 | 22.7 (2) | 140 |
| 2 | 32.0 (1) | 328 | 33.0 (1) | 398 |
| 3 | 39.6 (1) | 642 | 37.5 (1) | 536 |
| 4 | 45.5 (1) | 1290 | - | - |
| 5 | 55.6 ± 32 (8) | 1895 ± 354 | 58.0 ± 22 (10) | 2381 ± 309 |
| 6 | 64.4 ± 27 (21) | 3222 ± 682 | 63.5 ± 23 (12) | 3159 ± 806 |
| 7 | 72.1 ± 25 (30) | 4836 ± 761 | 71.9 ± 21 (16) | 4093 ± 807 |
| 8 | 79.5 ± 27 (52) | 7301 ± 1445 | 79.5 ± 22 (11) | 6266 ± 1240 |
| 9 | 85.4 ± 6 (14) | 8919 ± 882 | 86.6 (1) | 7706 |
| 10 | 87.4 ± 9 (10) | 9630 ± 2163 | - | - |
| 11 | 89.7 ± 4 (9) | 10,712 ± 849 | - | - |
| 12 | 91.3 ± 6 (8) | 10,668 ± 1185 | - | - |
| 13 | 92.3 ± 1 (3) | 10,520 ± 1091 | - | - |
| 14 | 93.9 ± 4 (3) | 12,170 ± 1517 | - | - |
| 15 | 95.9 (1) | 11,586 | - | - |
| 16 | 96.9 (1) | 12,672 | - | - |
| 17 | 97.5 ± 6 (3) | 13,433 ± 175 | - | - |
| 18 | 99.3 (1) | 13,100 | - | - |
| 19 | 99.8 (1) | 13,260 | - | - |
| Band Number | 0 | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|---|
| Back-calculated DW | 17.8 | 45.1 | 51.7 | 57.1 | 61.6 | |
| S.D. | ||||||
| N | ||||||
| Observed DW | 21.4 | 32.0 | 39.6 | 45.5 | 55.6 | |
| S.D. | 5 | 32 | ||||
| n | 2 | 1 | 1 | 1 | 8 |
| Species | Sex | n | Observed Max DW (cm) | Observed Max Age (Years) | DW∞ (cm) | t0 (Years) | k | Locality | Reference |
|---|---|---|---|---|---|---|---|---|---|
| R. bonasus | F | 40 | 107 | 13 | 125 | −3.764 | 0.119 | Chesapeake Bay (USA) | Smith and Merriner [31] |
| M | 51 | 98.1 | 8 | 119.2 | −3.699 | 0.126 | |||
| R. bonasus | F | 121 | 102.5 | 18+ | 123.8 * | −5.4799 * | 0.0746 | Northern Gulf of Mexico | Neer and Thompson [32] |
| M | 106 | 96 | 16+ | ||||||
| R. bonasus | F | 260 | 110.5 | 21 | 106.3 | −2.64 | 0.1931 | Chesapeake Bay (USA) | Fisher et al. [33] |
| M | 140 | 98 | 18 | 97.1 | −2.14 | 0.2741 | |||
| R. marginata | F | 170 | 99.8 | 19 | 178.6 | −2.082 | 0.122 | Iskenderun Bay (Turkey) | Present study |
| M | 54 | 86.6 | 9 | na | na | na |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Başusta, N.; Başusta, A.; Çiçek, E.; Cicia, A.M.; Sulikowski, J.A. First Estimates of Age and Growth of the Lusitanian Cownose Ray (Rhinoptera marginata) from the Mediterranean Sea. J. Mar. Sci. Eng. 2022, 10, 685. https://doi.org/10.3390/jmse10050685
Başusta N, Başusta A, Çiçek E, Cicia AM, Sulikowski JA. First Estimates of Age and Growth of the Lusitanian Cownose Ray (Rhinoptera marginata) from the Mediterranean Sea. Journal of Marine Science and Engineering. 2022; 10(5):685. https://doi.org/10.3390/jmse10050685
Chicago/Turabian StyleBaşusta, Nuri, Asiye Başusta, Erdoğan Çiçek, Angela M. Cicia, and James A. Sulikowski. 2022. "First Estimates of Age and Growth of the Lusitanian Cownose Ray (Rhinoptera marginata) from the Mediterranean Sea" Journal of Marine Science and Engineering 10, no. 5: 685. https://doi.org/10.3390/jmse10050685