

Relationship between Phenotypes and Chemical Profiling of Boesenbergia rotunda Collected from Different Habitats of Northern Thailand

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagent and Chemicals
2.2. Plant Materials
2.3. Phenotypic Characterization
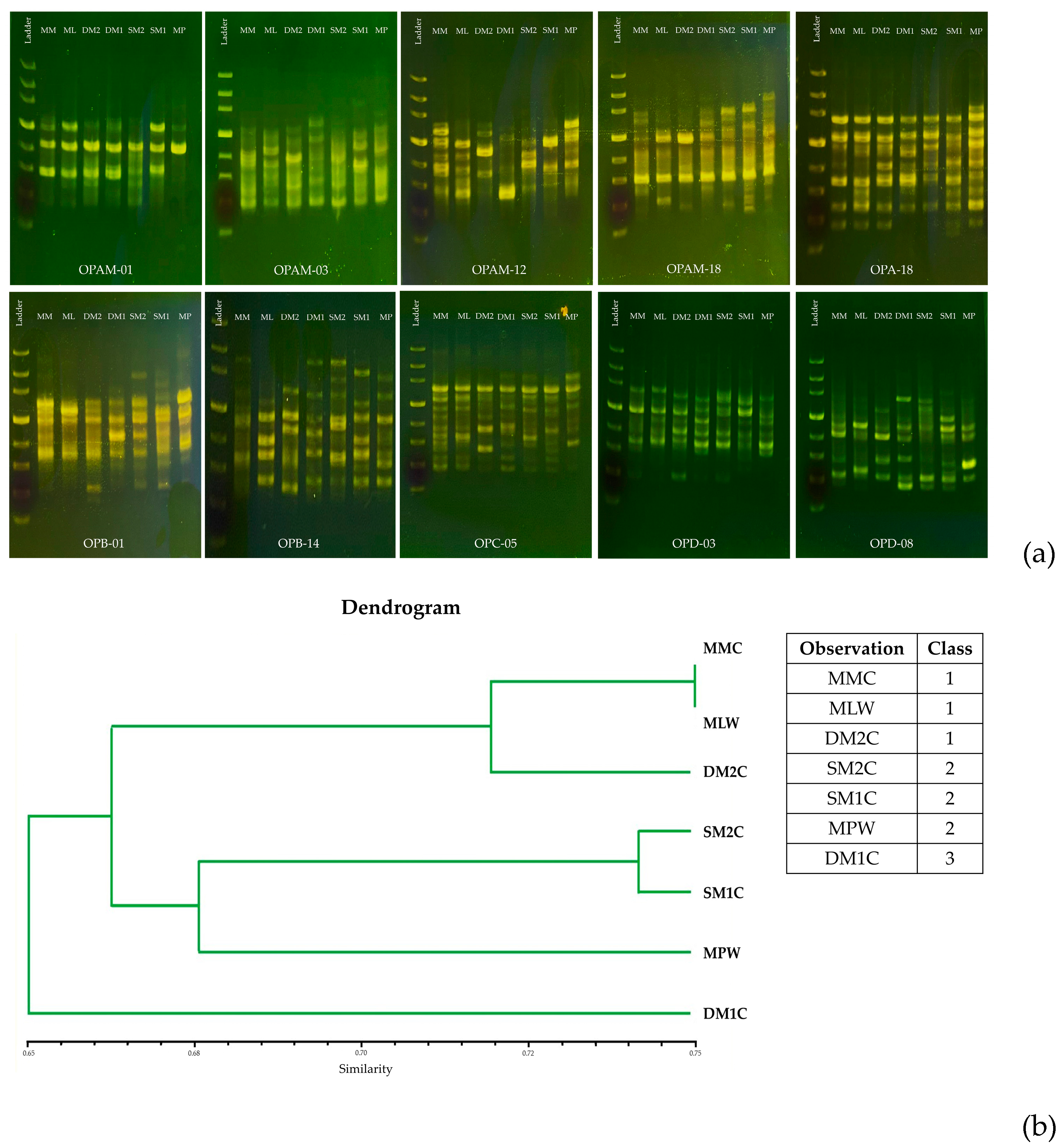
2.4. Genetic Variation Analysis of B. rotunda
2.4.1. DNA Extraction
2.4.2. RAPD-PCR
2.5. Phytochemical Analyses
2.5.1. Rhizome Extraction of B. rotunda
2.5.2. Total Phenolic Content
2.5.3. Total Flavonoid Content
2.5.4. Antioxidant Potential
DPPH Radical Scavenging Activity
ABTS•+ Radical Cation Activity
2.5.5. Analyses of Pinostrobin and Panduratin A by High-Performance Liquid Chromatography (HPLC)
2.5.6. Non-Volatile Metabolite Profiling Using Liquid Chromatography-Mass Spectrometer (LC-MS)
2.5.7. Volatile Metabolite Profiling Using Pyrolysis Gas Chromatography Mass Spectrometry (Py-GC-MS)
2.6. Statistical Analysis
3. Results and Discussion
3.1. Morphology
3.2. Genetic Variation of B. rotunda
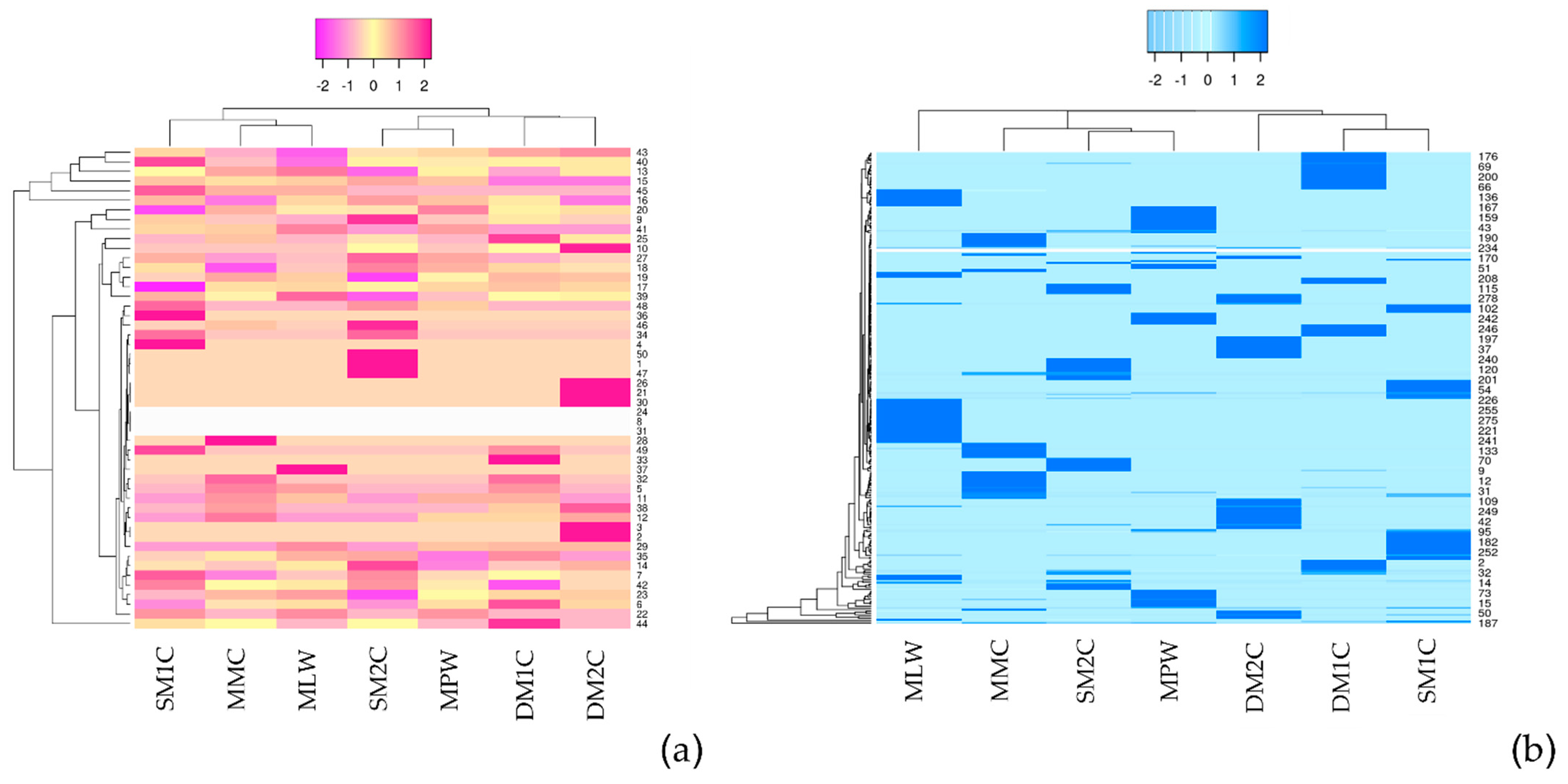
3.3. Phytochemical Compositions of B. rotunda
3.4. Metabolite Profiling of B. rotunda
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tewtrakul, S.; Subhadhirasakul, S. Anti-allergic activity of some selected plants in the Zingiberaceae family. J. Ethnopharmacol. 2007, 109, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Cheenpracha, S.; Karalai, C.; Ponglimanont, C.; Subhadhirasakul, S.; Tewtrakul, S. Anti-HIV-1 protease activity of compounds from Boesenbergia pandurata. Bioorg. Med. Chem. 2006, 14, 1710–1714. [Google Scholar] [CrossRef] [PubMed]
- Yuliana, N.D.; Budijanto, S.; Verpoorte, R.; Choi, Y.H.J. NMR metabolomics for identification of adenosine A1 receptor binding compounds from Boesenbergia rotunda rhizomes extract. J. Ethnopharmacol. 2013, 150, 95–99. [Google Scholar] [CrossRef]
- Chomchalow, N.; Bansiddhi, J.; MacBaine, C. Amazing Thai Medicinal Plants; Horticultural Research Institute Department of Agricultural, and Horticultural Science Society of Thailand: Bangkok, Thailand, 2003. [Google Scholar]
- Chuakul, W.; Boonpleng, A. Ethnomedical Uses of Thai Zingiberaceous Plant (1); Mahidol University: Bangkok, Thailand, 2003. [Google Scholar]
- Mood, J.; Tanaka, N.; Aung, M.; Murata, J. The genus Boesenbergia (Zingiberaceae) in Myanmar with two new records. Gard. Bull. Singap. 2016, 68, 299–318. [Google Scholar] [CrossRef]
- Kanjanasirirat, P.; Suksatu, A.; Manopwisedjaroen, S.; Munyoo, B.; Tuchinda, P.; Jearawuttanakul, K.; Seemakhan, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Rangkasenee, N. High-content screening of Thai medicinal plants reveals Boesenbergia rotunda extract and its component Panduratin A as anti-SARS-CoV-2 agents. Sci. Rep. 2020, 10, 19963. [Google Scholar] [CrossRef]
- Jing, L.J.; Mohamed, M.; Rahmat, A.; Bakar, M.F.A. Phytochemicals, antioxidant properties and anticancer investigations of the different parts of several gingers species (Boesenbergia rotunda, Boesenbergia pulchella var attenuata and Boesenbergia armeniaca). J. Med. Plants 2010, 4, 27–32. [Google Scholar]
- Ng, T.L.M.; Karim, R.; Tan, Y.S.; Teh, H.F.; Danial, A.D.; Ho, L.S.; Khalid, N.; Appleton, D.R.; Harikrishna, J.A. Amino acid and secondary metabolite production in embryogenic and non-embryogenic callus of fingerroot ginger (Boesenbergia rotunda). PLoS ONE 2016, 11, e0156714. [Google Scholar] [CrossRef]
- Eng-Chong, T.; Yean-Kee, L.; Chin-Fei, C.; Choon-Han, H.; Sher-Ming, W.; Li-Ping, C.T.; Gen-Teck, F.; Khalid, N.; Abd Rahman, N.; Karsani, S.A.J.E.-B.C.; et al. Boesenbergia rotunda: From ethnomedicine to drug discovery. Evid.-Based Complement. Altern. Med. 2012, 2012, 473637. [Google Scholar] [CrossRef]
- Bhamarapravati, S.; Juthapruth, S.; Mahachai, W.; Mahady, G.J.S. Antibacterial activity of Boesenbergia rotunda (L.) Mansf. and Myristica fragrans Houtt. against Helicobacter pylori. Songklanakarin J. Sci. Technol. 2006, 28, 157–163. [Google Scholar]
- Hwang, J.-K.; Chung, J.-Y.; Baek, N.-I.; Park, J.-H. Isopanduratin A from Kaempferia pandurata as an active antibacterial agent against cariogenic Streptococcus mutans. Int. J. Antimicrob. Agents 2004, 23, 377–381. [Google Scholar] [CrossRef]
- Jaipetch, T.; Kanghae, S.; Pancharoen, O.; Patrick, V.; Reutrakul, V.; Tuntiwachwuttikul, P.; White, A. Constituents of Boesenbergia pandurata (syn. Kaempferia pandurata): Isolation, crystal structure and synthesis of (±)-Boesenbergin A. Aust. J. Chem. 1982, 35, 351–361. [Google Scholar] [CrossRef]
- Noor Diyana, M.M. Flavonoid Pathway Gene Discoveries in Boesenbergia Rotunda through RNA-SEQ Transcriptome Profiling of Cell Suspension Cultures in Response to Phenylalanine/Noor Diyana Md Mustafa. Doctoral Dissertation, University of Malaya, Kuala Lumpur, Malaysia, 2017. [Google Scholar]
- Kirana, C.; Record, I.R.; McIntosh, G.H.; Jones, G.P. Screening for antitumor activity of 11 species of Indonesian zingiberaceae using human MCF-7 and HT-29 cancer cells. Pharm. Biol. 2003, 41, 271–276. [Google Scholar] [CrossRef]
- Kiat, T.S.; Pippen, R.; Yusof, R.; Ibrahim, H.; Khalid, N.; Abd Rahman, N. Inhibitory activity of cyclohexenyl chalcone derivatives and flavonoids of fingerroot, Boesenbergia rotunda (L.), towards dengue-2 virus NS3 protease. Bioorga. Med. Chem. Lett. 2006, 16, 3337–3340. [Google Scholar] [CrossRef]
- Watanaponpan, S.; Sairuksa, D.; Singhalert, R. White Galingale Cultivation for Promoting Community Economy of Farmers in Tha Maka District, Kanchanaburi Province. Interdiscip. Acad. Res. J. 2022, 2, 499–512. [Google Scholar]
- Prathepha, P. Screening of random primer to evaluate DNA diversity in Thai Curcuma using random amplified polymorphic DNAs. Warasan Songkhlanakarin 2000, 22, 7–13. [Google Scholar]
- Vanijajiva, O.; Sirirugsa, P.; Suvachittanont, W. Confirmation of relationships among Boesenbergia (Zingiberaceae) and related genera by RAPD. Biochem. Syst. Ecol. 2005, 33, 159–170. [Google Scholar] [CrossRef]
- Sangta, J.; Wongkaew, M.; Tangpao, T.; Withee, P.; Haituk, S.; Arjin, C.; Sringarm, K.; Hongsibsong, S.; Sutan, K.; Pusadee, T.J.P. Recovery of polyphenolic fraction from arabica coffee pulp and its antifungal applications. Plants 2021, 10, 1422. [Google Scholar] [CrossRef]
- Sunanta, P.; Chung, H.H.; Kunasakdakul, K.; Ruksiriwanich, W.; Jantrawut, P.; Hongsibsong, S.; Sommano, S.R. Genomic relationship and physiochemical properties among raw materials used for Thai black garlic processing. Food Sci. Nutr. 2020, 8, 4534–4545. [Google Scholar] [CrossRef]
- Ruttanapattanakul, J.; Wikan, N.; Okonogi, S.; Takuathung, M.N.; Buacheen, P.; Pitchakarn, P.; Potikanond, S.; Nimlamool, W. Boesenbergia rotunda extract accelerates human keratinocyte proliferation through activating ERK1/2 and PI3K/Akt kinases. Biomed. Pharmacother. 2021, 133, 111002. [Google Scholar] [CrossRef]
- Jirakiattikul, Y.; Rithichai, P.; Prachai, R.; Itharat, A. Elicitation enhancement of bioactive compound accumulation and antioxidant activity in shoot cultures of Boesenbergia rotunda L. Agric. Nat. Resour. 2021, 55, 456–463. [Google Scholar]
- Picó, Y.; Barceló, D. Pyrolysis gas chromatography-mass spectrometry in environmental analysis: Focus on organic matter and microplastics. TrAC Trends Anal. Chem. 2020, 130, 115964. [Google Scholar] [CrossRef]
- Chiavari, G.; Galletti, G.C.; Lanterna, G.; Mazzeo, R. The potential of pyrolysis—Gas chromatography/mass spectrometry in the recognition of ancient painting media. J. Anal. Appl. Pyrolysis 1993, 24, 227–242. [Google Scholar] [CrossRef]
- Fakhrudin, N.; Wiyono, T.; Putra, A.R.; Nurrochmad, A.; Widyarini, S.J.T. The evaluation on anti-platelet and antithrombosis activities of Cinnamomum sintoc bark extract. Thai J. Pharm. Sci. 2020, 43, 219–226. [Google Scholar]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef]
- Buxton, D.R.; Fales, S.L. Plant environment and quality. Forage Qual. Eval. Util. 1994, 155–199. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Li, Y.-S.; Liao, P.-C.; Chang, C.-T.; Hwang, S.-Y. Pattern of Adaptive Divergence in Zingiber kawagoii Hayata (Zingiberaceae) along a Narrow Latitudinal Range. Plants 2022, 11, 2490. [Google Scholar] [CrossRef]
- Gao, B.; Yuan, L.; Tang, T.; Hou, J.; Pan, K.; Wei, N. The complete chloroplast genome sequence of Alpinia oxyphylla Miq. and comparison analysis within the Zingiberaceae family. PLoS ONE 2019, 14, e0218817. [Google Scholar] [CrossRef]
- Bresson, J.; Vasseur, F.; Dauzat, M.; Koch, G.; Granier, C.; Vile, D. Quantifying spatial heterogeneity of chlorophyll fluorescence during plant growth and in response to water stress. Plant Methods 2015, 11, 23. [Google Scholar] [CrossRef]
- Li, H.; Huang, M.; Tan, D.; Liao, Q.; Zou, Y.; Jiang, Y. Effects of soil moisture content on the growth and physiological status of ginger (Zingiber officinale Roscoe). Acta Physiol. Plant. 2018, 40, 125. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.-S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef]
- Hallauer, A.R. Evolution of plant breeding. Crop Breed. Appl. Biotechnol. 2011, 11, 197–206. [Google Scholar] [CrossRef]
- Techaprasan, J.; Ngamriabsakul, C.; Klinbunga, S.; Chusacultanachai, S.; Jenjittikul, T. Genetic variation and species identification of Thai Boesenbergia (Zingiberaceae) analyzed by chloroplast DNA polymorphism. BMB Rep. 2006, 39, 361–370. [Google Scholar] [CrossRef]
- Techaprasan, J.; Klinbunga, S.; Jenjittikul, T. Genetic relationships and species authentication of Boesenbergia (Zingiberaceae) in Thailand based on AFLP and SSCP analyses. Biochem. Syst. Ecol. 2008, 36, 408–416. [Google Scholar] [CrossRef]
- Ebrahimi, F.; Mohammadi-Nejad, G.; Baghizadeh, A.; Abdolinejad, M. Genetic diversity evaluation of rapeseed genotypes (Brassica napus L.) based on phenotypic traits and random amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol. 2011, 10, 17391–17398. [Google Scholar] [CrossRef]
- Theanphong, O.; Mingvanish, W.; Jenjittikul, T.; Rungsihirunrat, K. Genetic variation of Boesenbergia rotunda (L.) Mansf. from Thailand based on essential oil compositions and internal transcribed spacer sequences. Songklanakarin J. Sci. Technol. 2021, 43, 1620–1627. [Google Scholar]
- Taheri, S.; Teo, C.H.; Heslop-Harrison, J.S.; Schwarzacher, T.; Tan, Y.S.; Wee, W.Y.; Khalid, N.; Biswas, M.K.; Mutha, N.V.R.; Mohd-Yusuf, Y.; et al. Genome Assembly and Analysis of the Flavonoid and Phenylpropanoid Biosynthetic Pathways in Fingerroot Ginger (Boesenbergia rotunda). Int. J. Mol. Sci. 2022, 23, 7269. [Google Scholar] [CrossRef]
- Gurung, A.B.; Ali, M.A.; Al-Hemaid, F.; El-Zaidy, M.; Lee, J. In silico analyses of major active constituents of fingerroot (Boesenbergia rotunda) unveils inhibitory activities against SARS-CoV-2 main protease enzyme. Saudi J. Biol. Sci. 2022, 29, 65–74. [Google Scholar] [CrossRef]
- Chahyadi, A.; Hartati, R.; Wirasutisna, K.R.J.P.C. Boesenbergia pandurata Roxb., an Indonesian medicinal plant: Phytochemistry, biological activity, plant biotechnology. Procedia Chem. 2014, 13, 13–37. [Google Scholar] [CrossRef]
- Saah, S.; Siriwan, D.; Trisonthi, P. Biological activities of Boesenbergia rotunda parts and extracting solvents in promoting osteogenic differentiation of pre-osteoblasts. Food Biosci. 2021, 41, 101011. [Google Scholar] [CrossRef]
- Wong, S.M.; Salim, N.; Harikrishna, J.A.; Khalid, N. Highly efficient plant regeneration via somatic embryogenesis from cell suspension cultures of Boesenbergia rotunda. Cell. Dev. Biol.-Plant 2013, 49, 665–673. [Google Scholar] [CrossRef]
- Mohammed, I.A.; Akhtar, M.N.; Biau, F.J.; Tor, Y.S.; Zareen, S.; Binti Shahabudin, S.; Binti Abd Hamid, H.; Ul Haq, Z.; Khalil, R.; Khalaf, R.M. Isolation of cardamonin and pinostrobin chalcone from the rhizomes of Boesenbergia rotunda (L.) Mansf. and their cytotoxic effects on H-29 and MDA-MB-231 cancer cell lines. Nat. Prod. J. 2019, 9, 341–348. [Google Scholar] [CrossRef]
- Yap, A.L.C. Phytochemical and Hplc Profiling of Extracts from Fingerroot (Boesenbergia rotunda) Rhizomes. Doctoral Dissertation, Universiti Putra Malaysia, Selangor, Malaysia, 2008. [Google Scholar]
- Jaudan, A.; Sharma, S.; Malek, S.N.A.; Dixit, A.J.P.O. Induction of apoptosis by pinostrobin in human cervical cancer cells: Possible mechanism of action. PLoS ONE 2018, 13, e0191523. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Fu, L.; Yuan, X.; Liu, Z.J.M. Promoting effect of pinostrobin on the proliferation, differentiation, and mineralization of murine pre-osteoblastic MC3T3-E1 cells. Molecules 2017, 22, 1735. [Google Scholar] [CrossRef] [PubMed]
- Athapaththu, A.M.G.K.; Lee, K.T.; Kavinda, M.H.D.; Lee, S.; Kang, S.; Lee, M.-H.; Kang, C.-H.; Choi, Y.H.; Kim, G.-Y. Pinostrobin ameliorates lipopolysaccharide (LPS)-induced inflammation and endotoxemia by inhibiting LPS binding to the TLR4/MD2 complex. Biomed. Pharmacother. 2022, 156, 113874. [Google Scholar] [CrossRef]
- Christov, R.; Trusheva, B.; Popova, M.; Bankova, V.; Bertrand, M. Chemical composition of propolis from Canada, its antiradical activity and plant origin. Nat. Prod. Res. 2006, 20, 531–536. [Google Scholar] [CrossRef]
- Patel, N.K.; Bhutani, K.K.J.P. Pinostrobin and Cajanus lactone isolated from Cajanus cajan (L.) leaves inhibits TNF-α and IL-1β production: In vitro and in vivo experimentation. Phytomedicine 2014, 21, 946–953. [Google Scholar] [CrossRef]
- Le Bail, J.-C.; Aubourg, L.; Habrioux, G. Effects of pinostrobin on estrogen metabolism and estrogen receptor transactivation. Cancer Lett. 2000, 156, 37–44. [Google Scholar] [CrossRef]
- Wu, N.; Kong, Y.; Zu, Y.; Fu, Y.; Liu, Z.; Meng, R.; Liu, X.; Efferth, T.J.P. Activity investigation of pinostrobin towards herpes simplex virus-1 as determined by atomic force microscopy. Phytomedicine 2011, 18, 110–118. [Google Scholar] [CrossRef]
- Xian, Y.-F.; Ip, S.-P.; Lin, Z.-X.; Mao, Q.-Q.; Su, Z.-R.; Lai, X.-P. Protective effects of pinostrobin on β-amyloid-induced neurotoxicity in PC12 cells. Cell. Mol. Neurobiol. 2012, 32, 1223–1230. [Google Scholar] [CrossRef]
- Patel, N.K.; Jaiswal, G.; Bhutani, K.K. A review on biological sources, chemistry and pharmacological activities of pinostrobin. Nat. Prod. Res. 2016, 30, 2017–2027. [Google Scholar] [CrossRef]
- Rukayadi, Y.; Han, S.; Yong, D.; Hwang, J.-K. In vitro antibacterial activity of panduratin A against enterococci clinical isolates. Biol. Pharm. Bull. 2010, 33, 1489–1493. [Google Scholar] [CrossRef]
- Kirana, C.; Jones, G.P.; Record, I.R.; McIntosh, G.H.J. Anticancer properties of panduratin A isolated from Boesenbergia pandurata (Zingiberaceae). J. Nat. Med. 2007, 61, 131–137. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental stress and secondary metabolites in plants: An overview. Plant Metab. Regul. Environ. Stress 2018, 153–167. [Google Scholar]
- Borges, C.V.; Minatel, I.O.; Gomez-Gomez, H.A.; Lima, G.P.P. Medicinal plants: Influence of environmental factors on the content of secondary metabolites. In Medicinal Plants and Environmental Challenges; Springer: Berlin/Heidelberg, Germany, 2017; pp. 259–277. [Google Scholar]
- Dey, M.; Kalia, S.; Ghosh, S.; Guha-Mukherjee, S. Biochemical and molecular basis of differentiation in plant tissue culture. Curr. Sci. 1998, 74, 591–596. [Google Scholar]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q.J.M. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Cultivated Area/GPS Coordinates | Habitat | Survivor Rate (%) | Leaf | Stems | Above Ground Part | ||
|---|---|---|---|---|---|---|---|---|
| Width (cm) | Length (cm) | Width (cm) | Height (cm) | |||||
| MMC | Mae Taeng, Chiang Mai/19.05992, 98.83022 | cultivation type | 100 | 8.03 ± 1.68 a | 25.43 ± 5.00 a | 1.45 ± 0.40 a | 51.75 ± 10.17 a |  |
| MLW | Mae Tha, Lamphun/18.63397, 99.04039 | wild type | 50 | 5.57 ± 1.63 b | 15.89 ± 5.25 bc | 1.08 ± 0.41 cd | 30.46 ± 10.45 c |  |
| DM1C | Chiang Dao, Chiang Mai/19.30270, 98.96640 | cultivation type | 95 | 7.28 ± 1.95 a | 17.71 ± 4.81 b | 1.29 ± 0.32 ab | 33.97 ± 11.35 bc |  |
| DM2C | Chiang Dao, Chiang Mai/19.51182, 99.09041 | cultivation type | 80 | 7.25 ± 1.65 a | 23.23 ± 4.57 b | 1.33 ± 0.32 ab | 44.43 ± 10.02 c |  |
| SM1C | San Pa Tong1, Chiang Mai/18.62599, 98.88333 | cultivation type | 50 | 5.74 ± 1.82 b | 17.28 ± 5.53 b | 0.96 ± 0.27 bc | 33.78 ± 12.78 c |  |
| SM2C | San Pa Tong2, Chiang Mai/18.62599, 98.88367 | cultivation type | 100 | 7.82 ± 2.04 a | 19.17 ± 5.52 a | 1.44 ± 0.45 a | 36.59 ± 12.72 ab |  |
| MPW | Mae Moh, Lampang/18.42797, 99.82055 | wild type | 45 | 4.30 ± 1.02 b | 12.42 ± 3.01 c | 0.67 ± 0.28 d | 21.40 ± 3.60 d |  |
| Sample | Mother Rhizomes | Fingers | Rhizome | ||||
|---|---|---|---|---|---|---|---|
| Weight (g) | Width (cm) | Length (cm) | Weight (g) | Width (cm) | Length (cm) | ||
| MMC | 8.24 ± 2.66 ab | 14.49 ± 0.56 a | 17.61 ± 2.70 ns | 47.07 ± 31.69 a | 12.69 ± 0.79 a | 5.86 ± 0.97 ns |  |
| MLW | 4.36 ± 2.84 b | 12.53 ± 0.28 b | 13.90 ± 1.03 ns | 20.79 ± 1.75 ab | 11.44 ± 3.24 ab | 6.50 ± 1.59 ns |  |
| DM1C | 14.16 ± 5.61 a | 14.07 ± 0.89 a | 17.87 ± 2.60 ns | 37.53 ± 25.23 ab | 13.47 ± 0.39 a | 6.49 ± 2.32 ns |  |
| DM2C | 3.23 ± 1.38 b | 12.13 ± 0.62 bc | 16.98 ± 3.43 ns | 23.51 ± 2.45 ab | 12.89 ± 2.49 a | 6.92 ± 1.04 ns |  |
| SM1C | 2.84 ± 2.25 b | 10.06 ± 0.62 d | 13.71 ± 2.94 ns | 19.30 ± 3.11 ab | 12.73 ± 0.89 a | 5.94 ± 1.08 ns |  |
| SM2C | 15.42 ± 7.63 a | 13.26 ± 0.39 ab | 17.84 ± 1.33 ns | 34.83 ± 24.39 ab | 13.96 ± 0.60 a | 6.83 ± 0.68 ns |  |
| MPW | 2.99 ± 1.05 b | 11.18 ± 1.14 cd | 16.97 ± 4.45 ns | 4.87 ± 1.87 b | 9.31 ± 2.06 b | 5.32 ± 5.10 ns |  |
| Sample | Pinostrobin (mg/gdried sample) | Panduratin A (mg/gdried sample) | Total Phenolic (mggallic/gdried sample) | Total Flavonoids (mgcatechin/gdried sample) | DPPH (%) | ABTS (%) |
|---|---|---|---|---|---|---|
| MMC | 27.09 ± 6.97 bc | 4.90 ± 1.16 bc | 20.80 ± 2.64 ab | 73.71 ± 17.04 b | 30.76 ± 5.88 a | 92.25 ± 0.52 b |
| MLW | 30.05 ± 7.55 bc | 5.83 ± 0.58 ab | 19.69 ± 0.34 bc | 50.00 ± 10.82 c | 24.99 ± 2.33 ab | 92.59 ± 1.27 ab |
| DM1C | 35.38 ± 10.43 ab | 6.06 ± 1.50 ab | 16.97 ± 2.12 c | 57.46 ± 13.36 bc | 24.57 ± 2.73 ab | 93.28 ± 0.43 ab |
| DM2C | 27.56 ± 8.30 bc | 5.04 ± 1.49 bc | 23.79 ± 1.15 a | 65.62 ± 2.75 bc | 27.71 ± 3.47 a | 93.82 ± 0.39 a |
| SM1C | 14.41 ± 2.43 c | 3.08 ± 0.81 c | 16.52 ± 1.99 c | 63.23 ± 3.78 bc | 14.07 ± 5.45 b | 93.08 ± 1.44 ab |
| SM2C | 43.27 ± 5.15 a | 7.49 ± 1.11 a | 19.22 ± 1.92 bc | 54.99 ± 7.45 bc | 26.44 ± 4.60 a | 92.69 ± 1.39 ab |
| MPW | 17.27 ± 6.24 c | 3.10 ± 0.69 c | 22.88 ± 2.07 ab | 95.12 ± 31.00 a | 23.65 ± 2.68 ab | 93.96 ± 0.53 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomya, S.; Wongkaew, M.; Bundithya, W.; Lumsangkul, C.; Jantrawut, P.; Sommano, S.R. Relationship between Phenotypes and Chemical Profiling of Boesenbergia rotunda Collected from Different Habitats of Northern Thailand. Agronomy 2023, 13, 1435. https://doi.org/10.3390/agronomy13061435
Thomya S, Wongkaew M, Bundithya W, Lumsangkul C, Jantrawut P, Sommano SR. Relationship between Phenotypes and Chemical Profiling of Boesenbergia rotunda Collected from Different Habitats of Northern Thailand. Agronomy. 2023; 13(6):1435. https://doi.org/10.3390/agronomy13061435
Chicago/Turabian StyleThomya, Sureerat, Malaiporn Wongkaew, Weenun Bundithya, Chompunut Lumsangkul, Pensak Jantrawut, and Sarana Rose Sommano. 2023. "Relationship between Phenotypes and Chemical Profiling of Boesenbergia rotunda Collected from Different Habitats of Northern Thailand" Agronomy 13, no. 6: 1435. https://doi.org/10.3390/agronomy13061435