
Response of Temperate Leymus chinensis Meadow Steppe Plant Community Composition, Biomass Allocation, and Species Diversity to Nitrogen and Phosphorus Addition
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
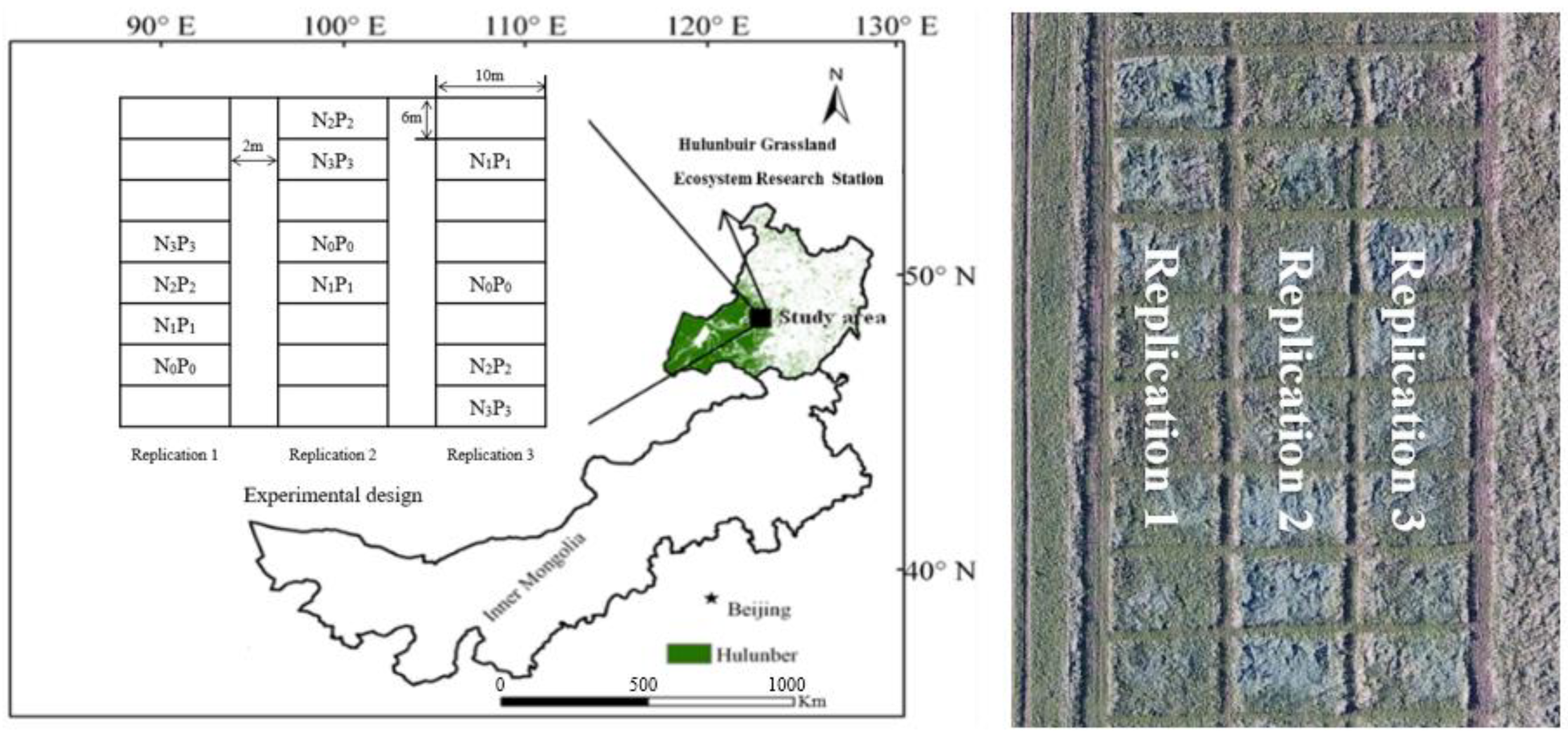
2.1. Overview of the Experimental Site
2.2. Fertilizer Application Trial Design
2.3. Research Methodology
2.3.1. Data Collection
2.3.2. Data Calculation and Processing
3. Results
3.1. Effect of N and P Addition on Plant Community Composition and Quantitative Characteristics
3.2. Effect of N and P Addition on Plant Biomass and Its Allocation
3.2.1. Effect of N and P Addition on Plant Community Biomass and Root-to-Shoot Ratio
3.2.2. Biomass of Plant Families in the Community after N and P Addition
3.3. Effect of N and P Addition on Community Species Diversity Indices
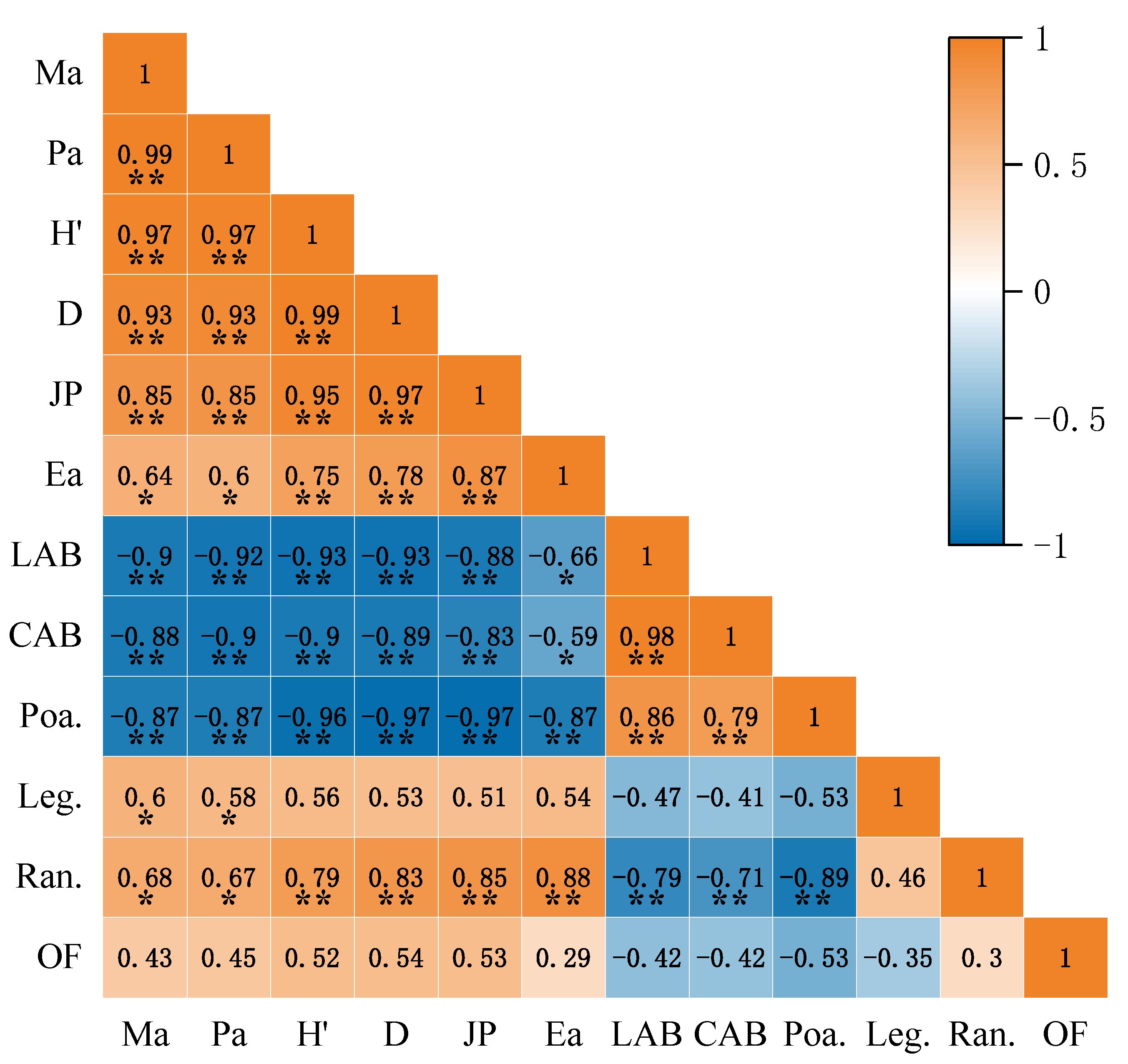
3.4. Effect of N and P Addition on the Correlation of Community Species Diversity Indices, Biomass, and Plant Functional Group Importance Values
4. Discussion
4.1. Effects of N and P Addition on the Species Composition of Communities and Their Quantitative Characteristics
4.2. Effects of N and P Addition on Grassland Biomass
4.3. Effects of N and P Addition on Plant Community Diversity
4.4. Correlation Analysis of Functional Groups, Biomass, and Diversity under Nitrogen and Phosphorus Addition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hao, L.; Sun, G.; Liu, Y.Q.; Gao, Z.Q.; He, J.J.; Shi, T.T.; Wu, B.J. Effects of precipitation on grassland ecosystem restoration under grazing exclusion in Inner Mongolia, China. Landsc. Ecol. 2014, 29, 1657–1673. [Google Scholar] [CrossRef]
- Dong, S.K.; Sherman, R. Enhancing the resilience of coupled human and natural systems of alpine rangelands on the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Akiyama, T.; Yokota, H.O.; Tsutsumi, M.; Yasuda, T.; Watanabe, O.; Wang, S.P. Quantifying grazing intensities using geographic information systems and satellite remote sensing in the Xilingol steppe region, Inner Mongolia, China. Agric. Ecosyst. Environ. 2005, 107, 83–93. [Google Scholar] [CrossRef]
- Dong, S.K.; Shang, Z.H.; Gao, J.X.; Boone, R.B. Enhancing sustainability of grassland ecosystems through ecological restoration and grazing management in an era of climate change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]
- Báez, S.; Fargione, J.; Moore, D.I.; Collins, S.L.; Gosz, J.R. Atmospheric nitrogen deposition in the northern Chihuahuan desert: Temporal trends and potential consequences. J. Arid Environ. 2007, 68, 640–651. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Li, M.Y.; Zhao, P.J.; Wu, L.B.; Chen, K. Effects of value perception, environmental regulation and their interaction on the improvement of herdsmen’s grassland ecological policy satisfaction. Int. J. Environ. Res. Public Health 2021, 18, 3078. [Google Scholar] [CrossRef]
- Wang, J.; Geng, Z.Z.; Xiao, H.F. Influence of Grassland Ecological Protection Subsidy-award Policies on Breeding Scale in China: An Empirical Study Based on Provincial Panel Data. J. PKU Aeronaut. Astronaut. Soc. Sci. 2018, 31, 84–93. [Google Scholar] [CrossRef]
- Basto, S.; Thompson, K.; Phoenix, G.; Sloan, V.; Leake, J.; Rees, M. Long-term nitrogen deposition depletes grassland seed banks. Nat. Commun. 2015, 6, 6185. [Google Scholar] [CrossRef] [Green Version]
- Virágh, K.; Tóth, T.; Somodi, I. Effect of slight vegetation degradation on soil properties in Brachypodium pinnatum grasslands. Plant Soil 2011, 345, 303–313. [Google Scholar] [CrossRef]
- Li, S.Y.; Verburg, P.H.; Lv, S.H.; Wu, J.L.; Li, X.B. Spatial analysis of the driving factors of grassland degradation under conditions of climate change and intensive use in Inner Mongolia, China. Reg. Environ. Chang. 2012, 12, 461–474. [Google Scholar] [CrossRef]
- Dlamini, P.; Chivenge, P.; Manson, A.; Chaplot, V. Land degradation impact on soil organic carbon and nitrogen stocks of subtropical humid grasslands in South Africa. Geoderma 2014, 235, 372–381. [Google Scholar] [CrossRef]
- Mekuria, W.; Veldkamp, E.; Haile, M.; Nyssen, J.; Muys, B.; Gebrehiwot, K. Effectiveness of exclosures to restore degraded soils as a result of overgrazing in Tigray, Ethiopia. J. Arid Environ. 2007, 69, 270–284. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Bernhardt, E.S. Biogeochemistry: An Analysis of Global Change; Academic Press: Waltham, MA, USA, 2013; pp. 1–672. ISBN 978-01-2385-874-0. [Google Scholar]
- Li, K.H.; Liu, X.J.; Song, L.; Gong, Y.M.; Lu, C.F.; Yue, P.; Tian, C.Y.; Zhang, F.S. Response of alpine grassland to elevated nitrogen deposition and water supply in China. Oecologia 2015, 177, 65–72. [Google Scholar] [CrossRef]
- Plassmann, K.; Edwards-Jones, G.; Jones, M.L.M. The effects of low levels of nitrogen deposition and grazing on dune grassland. Sci. Total Environ. 2009, 407, 1391–1404. [Google Scholar] [CrossRef]
- Gu, J.F.; Li, Z.K.; Mao, Y.Q.; Struik, P.C.; Zhang, H.; Liu, L.J.; Wang, Z.Q.; Yang, J.C. Roles of nitrogen and cytokinin signals in root and shoot communications in maximizing of plant productivity and their agronomic applications. Plant Sci. 2018, 274, 320–331. [Google Scholar] [CrossRef]
- Song, M.H.; Yu, F.H. Reduced compensatory effects explain the nitrogen-mediated reduction in stability of an alpine meadow on the Tibetan Plateau. New Phytol. 2015, 207, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Christopher, M.C.; Philip, E.M.; Frank, S.G.; Linda, H.P. Estimated losses of plant biodiversity in the United States from historical N deposition (1985–2010). Ecology 2013, 94, 1441–1448. [Google Scholar] [CrossRef]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M.; Belnap, J.; Brooks, M.L.; Cade, B.S.; Collins, S.L.; Geiser, L.H.; Gilliam, F.S.; et al. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef] [Green Version]
- Dai, J. Effects of Root Cutting and Fertilizer Application on Vegetation and Soil of Leymus Chinensis Meadow Mowing; Inner Mongolia Agricultural University: Huhehaote, China, 2016; pp. 13–29. [Google Scholar]
- Yan, R.R.; Xin, X.P.; Yan, Y.C.; Wang, X.; Zhang, B.H.; Yang, G.X.; Liu, S.M.; Deng, Y.; Li, L.H. Impacts of Differing Grazing Rates on Canopy Structure and Species Composition in Hulunber Meadow Steppe. Rangel. Ecol. Manag. 2015, 68, 54–64. [Google Scholar] [CrossRef]
- Zhang, Y.; Hou, L.L.; Yan, R.R.; Xin, X.P. Effects of grazing intensity on plant community characteristics and nutritional quality of meadow grassland. Chin. Agric. Sci. 2020, 53, 2550–2561. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Margalef, R. On certain unifying principles in ecology. Am. Nat. 1963, 97, 357–374. [Google Scholar] [CrossRef]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Ma, K.P.; Huang, J.H.; Yu, S.L. Study in species diversity, richness index, evenness index, species diversity index of vegetation community in Donglingshan Area in Beijing. Ecol. J. 1995, 15, 268–277. [Google Scholar] [CrossRef]
- Ma, K.P.; Liu, Y.M. Measure method of biocommunity’s diversity. Chin. Biodiv. 1994, 2, 231–239. [Google Scholar]
- Phoenix, G.K.; Hicks, W.K.; Cinderby, S.; Kuylenstierna, J.C.I.; Stock, W.D.; Dentener, F.J.; Giller, K.E.; Austin, A.T.; Lefroy, R.D.B.; Gimeno, B.S.; et al. Atmospheric nitrogen deposition in world biodiversity hotspots: The need for a greater global perspective in assessing N deposition impacts. Glob. Change Biol. 2006, 12, 470–476. [Google Scholar] [CrossRef]
- Reich, P.B.; Buschena, C.; Tjoelker, M.G.; Wrage, K.; Knops, J.; Tilman, D.; Machado, J.L. Variation in growth rate and ecophysiology among 34 grassland and savanna species under contrasting N supply: A test of functional group differences. New Phytol. 2003, 157, 617–631. [Google Scholar] [CrossRef]
- Suding, K.N.; Collins, S.L.; Gough, L.; Clark, C.; Cleland, E.E.; Gross, K.L.; Milchunas, D.G.; Pennings, S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. USA 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, W.D.; Gartner, J.R.; Holland, K.; Wiedermann, M. Nitrogen Critical Loads for Alpine Vegetation and Terrestrial Ecosystem Response: Are We There Yet? Ecol. Appl. 2006, 16, 1183–1193. [Google Scholar] [CrossRef]
- Eardly, B.D.; Hannaway, D.B.; Bottomley, P.J. Nitrogen Nutrition and Yield of Seedling Alfalfa as Affected by Ammonium Nitrate Fertilization1. Agron. J. 1985, 77, 57–62. [Google Scholar] [CrossRef]
- Ren, G.X.; Liu, J.F.; Xu, D.W.; Hong, W.; Zheng, S.Q.; Huang, Z.S. Analysis on classification and species diversity of Pinus taiwanensis community in Daiyun Mountain National Nature Reserve. J. Plant Resour. Environ. 2011, 20, 82–88. [Google Scholar] [CrossRef]
- Birhanu, L.; Bekele, T.; Tesfaw, B.; Demissew, S.H.M. Relationships between topographic factors, soil and plant communities in a dry Afromontane Forest Patches of Northwestern Ethiopia. PLoS ONE 2021, 16, e0247966. [Google Scholar] [CrossRef]
- Spehn, E.M.; Joshi, J.; Schmid, B.; Diemer, M.; Körner, C. Above-groud resource use increase with plant species richness in experiment grassland ecosystem. Funct. Ecol. 2000, 14, 326–337. [Google Scholar] [CrossRef]
- Jiang, Y.X.; Wang, B.S.; Zang, R.G.; Jin, J.H.; Liao, W.B. The Biodiversity and Its Formation Mechanism of Tropical Forests in Hainan Island; Science Press: Beijing, China, 2002; pp. 217–233. [Google Scholar]
- Feng, Y.Z. Species diversity and managed ecosystem stability. J. Appl. Ecol. 2003, 14, 853–857. [Google Scholar]
- Liu, Z.Y.; Baoyin, T.G.T.; Duan, J.J.; Yang, G.F.; Sun, J.; Li, X.L. Nutrient Characteristics in Relation to Plant Size of a Perennial Grass Under Grazing Exclusion in Degraded Grassland. Front. Plant Sci. 2018, 9, 295. [Google Scholar] [CrossRef]
- Zhou, X.L.; Guo, Z.; Zhang, P.F.; Du, G.Z. Shift in community functional composition following nitrogen fertilization in an alpine meadow through intraspecific trait variation and community composition change. Plant Soil 2018, 431, 289–302. [Google Scholar] [CrossRef]
- Schellberg, J.; Pontes, L.S. Plant functional traits and nutrient gradients on grassland. Grass Forage Sci. 2012, 67, 305–319. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef]
- Zhou, X.L.; Guo, Z.; Zhang, P.F.; Li, H.L.; Chu, C.J.; Li, X.L.; Du, G.Z. Different categories of biodiversity explain productivity variation after fertilization in a Tibetan alpine meadow community. Ecol. Evol. 2017, 7, 3464–3474. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.M.; Baoyin, T.G.T.; Li, Y.R.; Li, Y.H.; Chen, J.S. Analysis of the effect of root cutting method and nitrogen addition on the improvement of degraded Leymus chinensis grassland. Chin. J. Grassl. 2017, 39, 67–71. [Google Scholar] [CrossRef]
- Zong, N.; Shi, P.L. Enhanced community production rather than structure improvement under nitrogen and phosphorus addition in severely degraded alpine meadows. Sustainability 2019, 11, 2023. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Shi, P.L.; Zong, N.; Fu, G.; Shen, Z.X.; Zhang, X.Z.; Song, M. H, Climatic patterns modulate ecosystem and soil respiration responses to fertilization in an alpine meadow on the Tibetan Plateau, China. Ecol. Res. 2015, 30, 3–13. [Google Scholar] [CrossRef]
- Yang, X.X.; Ren, F.; Zhou, H.K.; He, J.S. Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau. J. Plant Ecol. 2014, 38, 159–166. [Google Scholar] [CrossRef]
- Li, C.Q.; Li, Q.; Zhao, L.; Zhao, X.Q. Responses of plant community biomass to nitrogen and phosphorus additions in natural and restored grasslands around Qinghai Lake Basin. J. Plant Ecol. 2016, 40, 1015–1027. [Google Scholar] [CrossRef] [Green Version]
- Harpole, W.S.; Ngai, J.T.; Cleland, E.E.; Seabloom, E.W.; Borer, E.T.; Bracken, M.E.S.; Elser, J.J.; Gruner, D.S.; Hillebrand, H.; Shurin, J.B.; et al. Nutrient colimitation of primary producer communities. Ecol. Lett. 2011, 14, 852–862. [Google Scholar] [CrossRef]
- Carnicer, J.; Sardans, J.; Stefanescu, C.; Ubach, A.; Bartrons, M.; Asensio, D.; Penuelas, J. Global biodiversity, stoichiometry and ecosystem function responses to human-induced C–N–P imbalances. J. Plant Physiol. 2015, 172, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Bracken, M.E.S.; Hillebrand, H.; Borer, E.T.; Seabloom, E.W.; Cebrian, J.; Cleland, E.E.; Elser, J.J.; Gruner, D.S.; Harpole, W.S.; Ngai, J.T.; et al. Signatures of nutrient limitation and colimitation: Responses of autotroph internal nutrient concentrations to nitrogen and phosphorus additions. Oikos 2015, 124, 113–121. [Google Scholar] [CrossRef]
- Wei, C.Z.; Yu, Q.; Bai, E.; Lü, X.T.; Li, Q.; Xia, J.Y.; Kardol, P.; Liang, W.J.; Wang, Z.W.; Han, X.G. Nitrogen deposition weakens plant–microbe interactions in grassland ecosystems. Glob. Chang. Biol. 2013, 19, 3688–3697. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.Y.; Wan, S.Q. Global response patterns of terrestrial plant species to nitrogen addition. New Phytol. 2008, 179, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Fornara, D.A.; Banin, L.; Crawley, M.J. Multinutrient vs. nitrogen-only effects on carbon sequestration in grassland soils. Glob. Chang. Biol. 2013, 19, 3848–3857. [Google Scholar] [CrossRef] [Green Version]
- Johnston, A.E.; Poulton, P.R. The role of phosphorus in crop production and soil fertility:150 years of field experiments at Rothamsted, United Kingdon. In Phosphate Fertilizers and the Environment: Proceedings of an International Workshop; Schultz, J.J., Ed.; International Fertilizer Development Center: Tampa, FL, USA, 1992; pp. 45–64. [Google Scholar]
- Palta, J.A.; Yang, J.C. Crop root system behaviour and yield. Field Crop. Res. 2014, 165, 1–149. [Google Scholar] [CrossRef]
- Mohd-Radzman, N.A.; Djordjevic, M.A.; Imin, N. Nitrogen modulation of legume root architecture signalling pathways involves phytohormones and small regulatory molecules. Front. Plant Sci. 2013, 4, 385. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.B.; Zhang, Y.L.; Turner, D.; Yin-Hua, D.; Dong-Sheng, W.; Qi-Rong, S. Root physiological and morphological characteristics of two rice cultivars with different nitrogen-use efficiency. Pedosphere 2010, 20, 446–455. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Khan, A.; Tan, D.K.Y.; Luo, H.H. Rational water and nitrogen management improves root growth, increases yield and maintains water use efficiency of cotton under mulch drip irrigation. Front. Plant Sci. 2017, 8, 912. [Google Scholar] [CrossRef]
- Gaudin, A.C.M.; Mcclymont, S.A.; Holmes, B.M.; Lyons, E.; Raizada, M.N. Novel temporal, fine-scale and growth variation phenotypes in roots of adult-stage maize (Zea mays L.) in response to low nitrogen stress. Plant Cell Environ. 2011, 34, 2122–2137. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Li, L.H.; Chen, X.; Tian, X.; Wang, X.K.; Luo, G.P. Biomass Allocation Patterns across China’s Terrestrial Biomes. PLoS ONE 2014, 9, e93566. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mi, G.H.; Chen, F.J.; Zhang, J.H.; Zhang, F.S. Response of root morphology to nitrate supply and its contribution to nitrogen accumulation in maize. J. Plant Nutr. 2005, 27, 2189–2202. [Google Scholar] [CrossRef]
- Bengtsson, J. Which species? What kind of diversity? Which ecosystem function? Some problems in studies of relations between biodiversity and ecosystem function. Appl. Soil Ecol. 1998, 10, 191–199. [Google Scholar] [CrossRef]
- Naeem, S.; Li, S. Biodiversity enhance ecosystem reliability. Nature 1997, 390, 507–509. [Google Scholar] [CrossRef]
- Walker, M.D.; Webber, P.J.; Arnold, E.H.; Ebert-May, D. Effects of interannual climate variation on aboveground phytomass in alpine vegetation. Ecology 1994, 75, 393–408. [Google Scholar] [CrossRef]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in european grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Naeem, S.; Pan, Q.M.; Huang, J.H.; Zhang, L.X.; Han, X.G. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. Glob. Change Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Tilman, D. Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol. Monogr. 1987, 57, 189–214. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Huston, M.; Smith, T. Plant Succession: Life History and Competition. Am. Nat. 1987, 130, 168–198. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and nutrients control grassland plant diversity via light limitation. Nature 2014, 508, 517–520. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.L.; Hautier, Y.; Borer, E.T.; Zhang, C.H.; Du, G.Z. Abundance-and functional-based mechanisms of plant diversity loss with fertilization in the presence and absence of herbivores. Oecologia 2015, 179, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Harpole, W.S.; Sullivan, L.L.; Lind, E.M.; Firn, J.; Adler, P.B.; Borer, E.T.; Chase, J.; Fay, P.A.; Hautier, Y.; Hillebrand, H.; et al. Addition of multiple limiting resources reduces grassland diversity. Nature 2016, 537, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.N.; Yang, B.; Li, M.X.; Xiao, R.Q.; Rao, K.Y.; Wang, J.Q.; Zhang, T.; Guo, J.X. Community Composition, Structure and Productivity in Response to Nitrogen and Phosphorus Additions in a Temperate Meadow. Sci. Total Environ. 2019, 654, 863–871. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Minden, V.; Scherber, C.; Cebrián, P.; Miguel, A.; Trinogga, J.; Trenkamp, A.; Mantilla-Contreras, J.; Lienin, P.; Kleyer, M. Consistent drivers of plant biodiversity across managed ecosystems. Philos. Trans. R. Soc. B 2016, 371, 1694. [Google Scholar] [CrossRef] [Green Version]
- Drescher, J.; Rembold, K.; Allen, K.; Beckschäfer, P.; Buchori, D.; Clough, Y.; Faust, H.; Fauzi, A.M.; Gunawan, D.; Hertel, D.; et al. Ecological and socioeconomic functions across tropical land use systems after rainforest conversion. Philos. Trans. R. Soc. B 2016, 371, 1694. [Google Scholar] [CrossRef] [Green Version]
- Lawton, J.H. What do species do in ecosystems? Oikos 1994, 71, 367–374. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
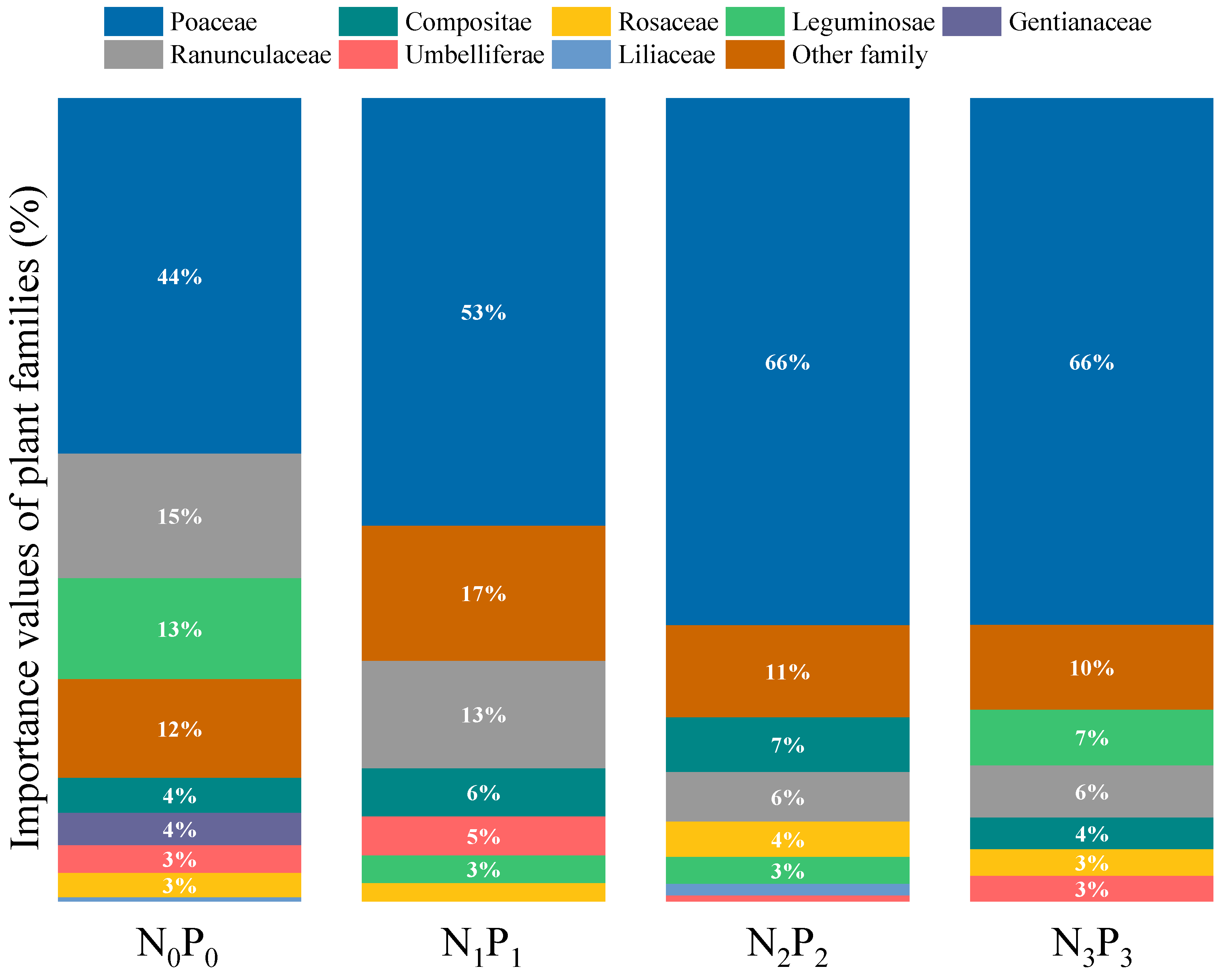{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Depth (cm) | TN (g·kg−1) | TP (g·kg−1) | TK (g·kg−1) | TC (g·kg−1) | pH |
|---|---|---|---|---|---|
| 0–10 | 3.28 | 0.51 | 22.62 | 37.79 | 6.97 |
| 10–20 | 2.77 | 0.50 | 24.23 | 29.33 | 7.01 |
| 20–30 | 2.12 | 0.45 | 22.38 | 21.67 | 7.13 |
| Family | Species | N0P0 | N1P1 | N2P2 | N3P3 |
|---|---|---|---|---|---|
| Poaceae | Leymus chinensis | 34.09 ± 8.69 | 50.92 ± 3.01 | 65.65 ± 1.98 | 65.62 ± 0.34 |
| Stipa baicalensis | 2.05 ± 2.05 | 0 | 0 | 0 | |
| Koeleria cristata | 0.44 ± 0.44 | 0.59 ± 0.59 | 0 | 0 | |
| Poa annua | 0.00 ± 0.00 | 1.32 ± 1.32 | 0 | 0 | |
| Achnatherum sibiricum | 6.09 ± 3.12 | 0 | 0 | 0 | |
| Cleistogenes squarrosa | 1.61 ± 0.09 | 0.43 ± 0.43 | 0 | 0 | |
| Compositae | Artemisia frigida | 0.42 ± 0.42 | 1.65 ± 0.95 | 1.12 ± 1.12 | 0 |
| Serratula centauroides | 1.74 ± 0.92 | 1.85 ± 1.85 | 1.39 ± 1.39 | 0 | |
| Lactuca sativa | 0 | 0 | 1.66 ± 1.66 | 0 | |
| Artemisia tanacetifolia | 2.25 ± 1.24 | 2.54 ± 1.32 | 2.71 ± 1.57 | 1.34 ± 1.34 | |
| Artemisia dracunculus | 0 | 0 | 0 | 1.52 ± 1.52 | |
| Heteropappus altaicus | 0 | 0 | 0 | 1.14 ± 1.14 | |
| Rosaceae | Potentilla tanacetifolia | 0.87 ± 0.87 | 0 | 3.08 ± 0.85 | 1.33 ± 1.33 |
| Potentilla acaulis | 0.21 ± 0.21 | 0 | 0 | 0 | |
| Potentilla bifurca | 1.99 ± 0.28 | 2.29 ± 1.30 | 1.38 ± 1.38 | 1.98 ± 1.98 | |
| Leguminosae | Vicia amoena | 7.30 ± 1.37 | 1.15 ± 1.15 | 1.25 ± 1.25 | 2.88 ± 2.88 |
| Thermopsis lanceolata | 0.96 ± 0.96 | 1.28 ± 1.28 | 0.88 ± 0.88 | 1.41 ± 1.41 | |
| Astragalus melilotoides | 4.35 ± 2.42 | 1.04 ± 1.04 | 1.23 ± 1.23 | 2.69 ± 1.36 | |
| Gentianaceae | Gentianopsis barbata | 0.91 ± 0.91 | 0 | 0 | 0 |
| Gentiana squarrosa | 0.94 ± 0.52 | 0 | 0 | 0 | |
| Lomatogonium micranthum | 2.17 ± 0.11 | 0 | 0 | 0 | |
| Ranunculaceae | Thalictrum squarrosum | 8.97 ± 4.89 | 10.27 ± 2.59 | 5.33 ± 0.41 | 6.46 ± 1.20 |
| Pulsatilla turczaninovii | 6.51 ± 4.08 | 3.12 ± 1.21 | 0.77 ± 0.77 | 0 | |
| Umbelliferae | Bupleurum scorzonerifolium | 3.42 ± 0.29 | 3.97 ± 0.21 | 0.76 ± 0.76 | 3.14 ± 1.60 |
| Saposhnikovia divaricata | 0 | 0.83 ± 0.83 | 0 | 0 | |
| Liliaceae | Allium ramosum | 0 | 0 | 1.38 ± 1.38 | 0 |
| Allium bidentatum | 0.48 ± 0.48 | 0 | 0 | 0 | |
| Other families | Carex duriuscula | 5.94 ± 1.66 | 5.13 ± 2.59 | 4.07 ± 2.19 | 3.19 ± 3.19 |
| Adenophora stricta | 1.67 ± 1.67 | 2.55 ± 1.36 | 0 | 1.84 ± 1.84 | |
| Dianthus chinensis | 0.81 ± 0.81 | 0 | 0 | 0 | |
| Galium verum | 2.95 ± 0.26 | 6.57 ± 1.56 | 4.37 ± 0.50 | 5.46 ± 0.56 | |
| Linaria vulgaris | 0 | 1.29 ± 1.29 | 0 | 0 | |
| Iris ventricosa | 0.84 ± 0.84 | 0 | 1.52 ± 1.52 | 0 | |
| Schizonepeta multifida | 0 | 1.22 ± 1.22 | 0 | 0 | |
| Dontostemon micranthus | 0 | 0 | 1.46 ± 1.46 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Xin, X.; Zhang, Y.; Wang, M.; Chen, S.; Yu, T.; Li, Y.; Yang, G.; Yan, R. Response of Temperate Leymus chinensis Meadow Steppe Plant Community Composition, Biomass Allocation, and Species Diversity to Nitrogen and Phosphorus Addition. Agronomy 2023, 13, 208. https://doi.org/10.3390/agronomy13010208
Zhang C, Xin X, Zhang Y, Wang M, Chen S, Yu T, Li Y, Yang G, Yan R. Response of Temperate Leymus chinensis Meadow Steppe Plant Community Composition, Biomass Allocation, and Species Diversity to Nitrogen and Phosphorus Addition. Agronomy. 2023; 13(1):208. https://doi.org/10.3390/agronomy13010208
Chicago/Turabian StyleZhang, Chu, Xiaoping Xin, Yu Zhang, Miao Wang, Sisi Chen, Tianqi Yu, Yingxin Li, Guixia Yang, and Ruirui Yan. 2023. "Response of Temperate Leymus chinensis Meadow Steppe Plant Community Composition, Biomass Allocation, and Species Diversity to Nitrogen and Phosphorus Addition" Agronomy 13, no. 1: 208. https://doi.org/10.3390/agronomy13010208