
Selection of Non-Crop Plant Mixes Informed by Arthropod-Plant Network Analyses for Multiple Ecosystem Services Delivery Towards Ecological Intensification of Agriculture

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Focal Observations of Plant—Arthropod Interactions
2.3. Data Analyses
3. Results
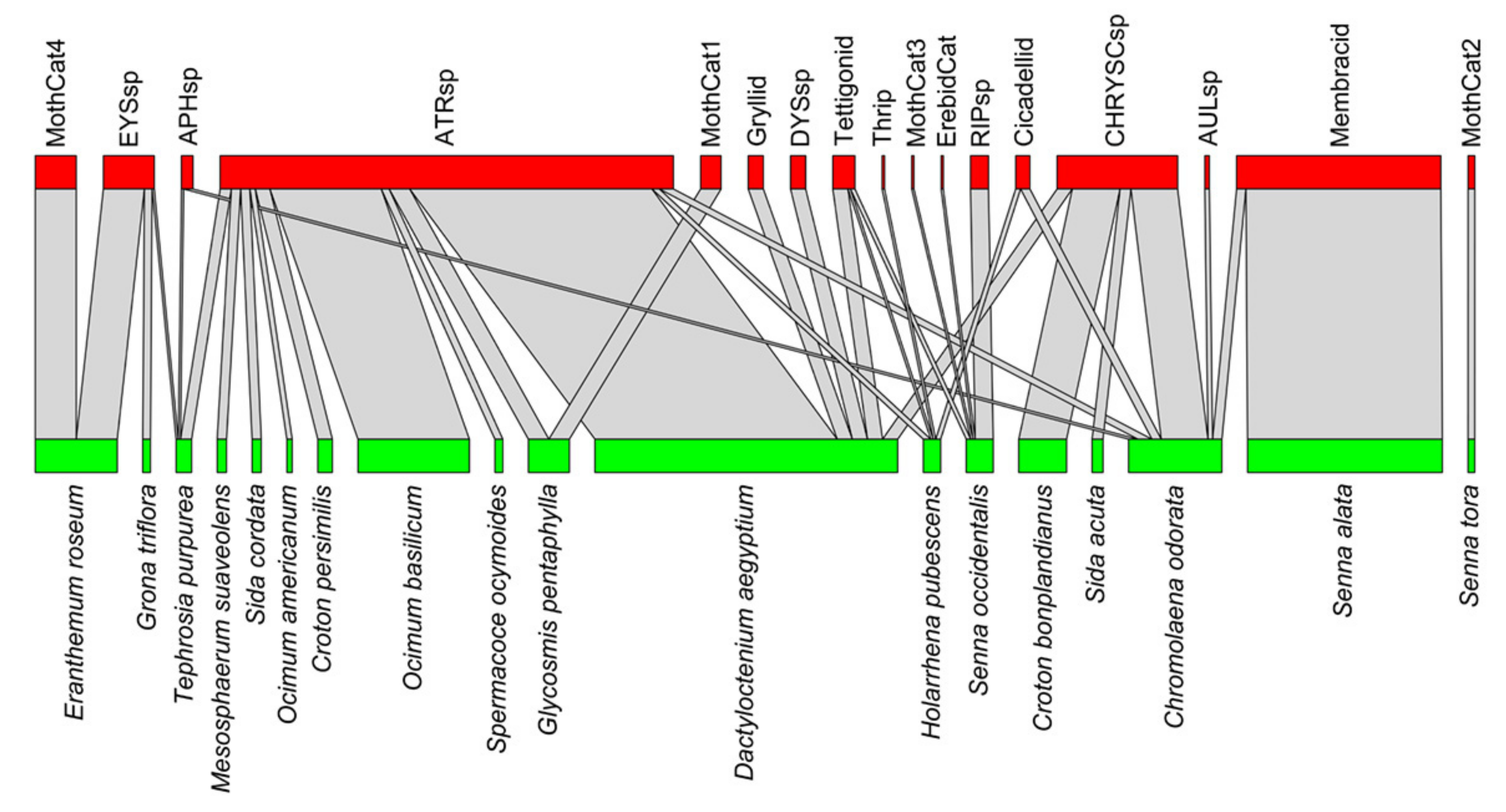
3.1. Comparison among Three Arthropod Communities Interacting with Non-Crop Plant Community
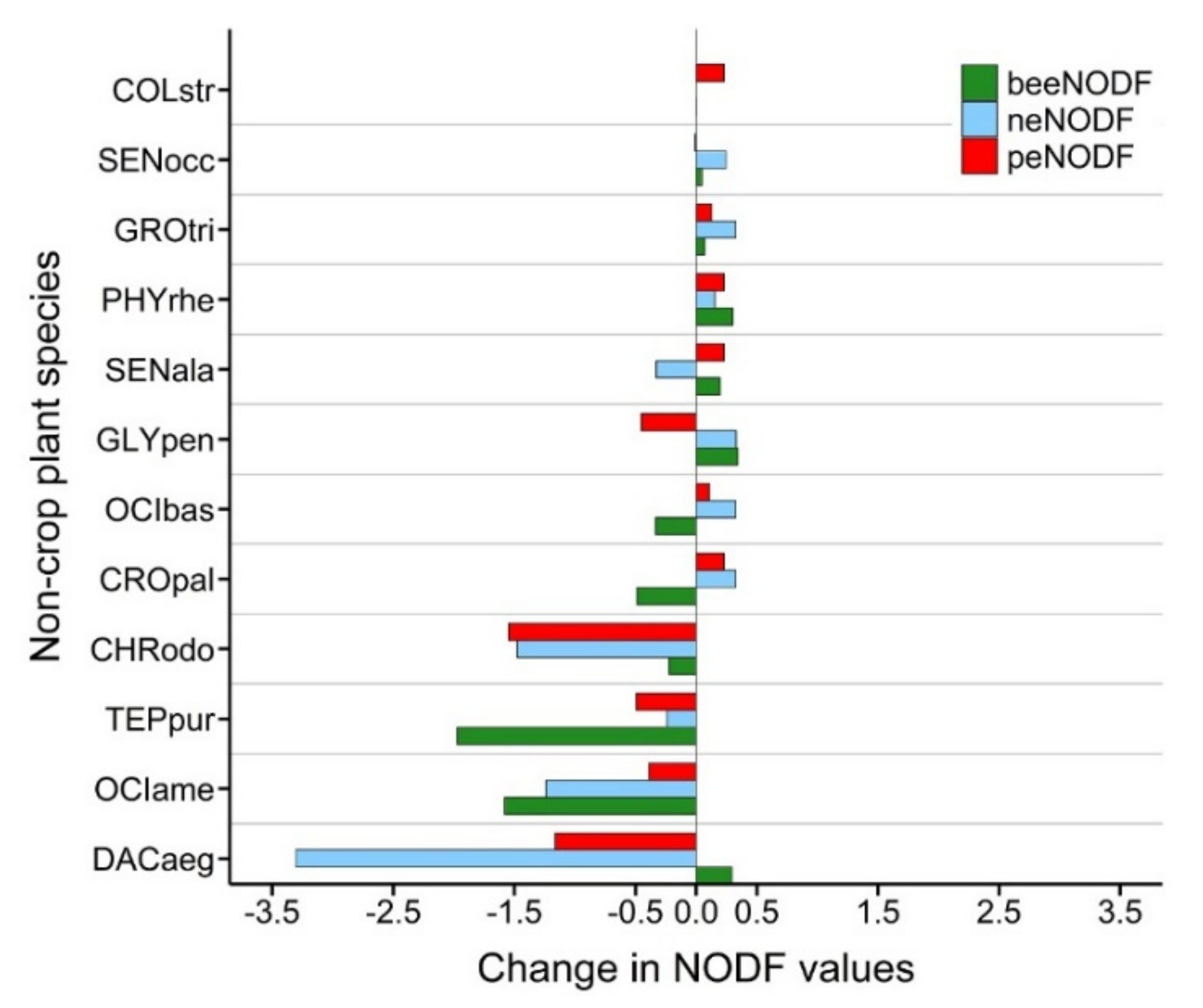
3.2. Contribution of Connector Non-Crop Species to the Nestedness of Networks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Bank. World Development Report 2008: Agriculture for Development; The World Bank: Washington, DC, USA, 2007. [Google Scholar]
- Crossman, N.D.; Connor, J.D.; Bryan, B.A.; Summers, D.M.; Ginnivan, J. Reconfiguring an irrigation landscape to improve provision of ecosystem services. Ecol. Econ. 2010, 69, 1031–1042. [Google Scholar] [CrossRef]
- Bolwig, S.; Tanner, A.N.; Riemann, P.; Redlingshöfer, B.; Zhang, Y. Reducing Consumer Food Waste Using Green and Digital Technologies. Copenhagen and Nairobi; UNEP, D. Partnership (United Nations Environment Programme): Copenhagen, Denmark, 2021. [Google Scholar]
- Dainese, M.; Martin, E.A.; Aizen, M.A.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A.; et al. A global synthesis reveals biodiversity-mediated benefits for crop production. Sci. Adv. 2019, 5, eaax0121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, P.H.; Wagner, D.L. Agricultural intensification and climate change are rapidly decreasing insect biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2002548117. [Google Scholar] [CrossRef] [PubMed]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V. Ecological intensification to mitigate impacts of conventional intensive land use on pollinators and pollination. Ecol. Lett. 2017, 20, 673–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Pérez-Méndez, N.; Garratt, M.P.; Gemmill-Herren, B.; Miguez, F.E.; Dicks, L.V. Policies for ecological intensification of crop production. Trends Ecol. Evol. 2019, 34, 282–286. [Google Scholar] [CrossRef] [Green Version]
- Tittonell, P. Ecological intensification of agriculture—Sustainable by nature. Curr. Opin. Environ. Sustain. 2014, 8, 53–61. [Google Scholar] [CrossRef]
- Williams, N.M.; Ward, K.L.; Pope, N.; Isaacs, R.; Wilson, J.; May, E.A.; Ellis, J.; Daniels, J.; Pence, A.; Ullmann, K.; et al. Native wildflower plantings support wild bee abundance and diversity in agricultural landscapes across the United States. Ecol. Appl. 2015, 25, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L.; Klatt, B.K.; Smith, H.G. Effects of Flower-Enriched Ecological Focus Areas on Functional Diversity Across Scales. Front. Ecol. Evol. 2021, 9, 288. [Google Scholar] [CrossRef]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.; Smith, B.M. Semi-natural habitats support biological control, pollination and soil conservation in Europe. A review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Montgomery, I.; Caruso, T.; Reid, N. Hedgerows as ecosystems: Service delivery, management, and restoration. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 81–102. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Zhou, W.; Lee, M.-B.; Goodale, E. The relationship between crop cover, crop heterogeneity and the diversity of herbaceous plants in uncultivated areas of a tropical agricultural landscape of southern China. Glob. Ecol. Conserv. 2018, 14, e00399. [Google Scholar] [CrossRef]
- DeFries, R.; Rosenzweig, C. Toward a whole-landscape approach for sustainable land use in the tropics. Proc. Natl. Acad. Sci. USA 2010, 107, 19627–19632. [Google Scholar] [CrossRef] [Green Version]
- Basu, P.; Parui, A.K.; Chatterjee, S.; Dutta, A.; Chakraborty, P.; Roberts, S.; Smith, B. Scale dependent drivers of wild bee diversity in tropical heterogeneous agricultural landscapes. Ecol. Evol. 2016, 6, 6983–6992. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Laha, S.; Chatterjee, S.; Das, A.; Smith, B.; Basu, P. Exploring the importance of floral resources and functional trait compatibility for maintaining bee fauna in tropical agricultural landscapes. J. Insect Conserv. 2020, 24, 431–443. [Google Scholar] [CrossRef]
- Hobbs, P.R.; Sayre, K.; Gupta, R. The role of conservation agriculture in sustainable agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 543–555. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Parker, J.E.; Crowder, D.W.; Eigenbrode, S.D.; Snyder, W.E. Trap crop diversity enhances crop yield. Agric. Ecosyst. Environ. 2016, 232, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Hokkanen, H.M. Trap cropping in pest management. Annu. Rev. Entomol. 1991, 36, 119–138. [Google Scholar] [CrossRef]
- Holden, M.H.; Ellner, S.P.; Lee, D.H.; Nyrop, J.P.; Sanderson, J.P. Designing an effective trap cropping strategy: The effects of attraction, retention and plant spatial distribution. J. Appl. Ecol. 2012, 49, 715–722. [Google Scholar] [CrossRef]
- Keyes, A.A.; McLaughlin, J.P.; Barner, A.K.; Dee, L.E. An ecological network approach to predict ecosystem service vulnerability to species losses. Nat. Commun. 2021, 12, 1586. [Google Scholar] [CrossRef]
- Bascompte, J.; Jordano, P.; Melian, C.J.; Olesen, J.M. The nested assembly of plant–animal mutualistic networks. Proc. Natl. Acad. Sci. USA 2003, 100, 9383–9387. [Google Scholar] [CrossRef] [Green Version]
- Cagua, E.F.; Wootton, K.L.; Stouffer, D.B. Keystoneness, centrality, and the structural controllability of ecological networks. J. Ecol. 2019, 107, 1779–1790. [Google Scholar] [CrossRef]
- González, A.M.M.; Dalsgaard, B.; Olesen, J.M. Centrality measures and the importance of generalist species in pollination networks. Ecol. Complex. 2010, 7, 36–43. [Google Scholar] [CrossRef]
- Mahapatra, N.; Marai, N.; Dhal, K.; Nayak, R.N.; Panigrahi, B.K.; Mallick, G.; Ranjit, M.; Kar, S.K.; Kerketta, A.S. Malaria outbreak in a non endemic tribal block of Balasore district, Orissa, India during summer season. Trop. Biomed. 2012, 29, 277–285. [Google Scholar]
- Sutherland, W.J. Ecological Census Techniques: A Handbook; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology; Wiley: Hoboken, NJ, USA, 2014. [Google Scholar]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Triplehorn, C.A.; Johnson, N.F.; Borror, D.J. Borror and DeLong’s Introduction to the Study of Insects; Brooks/Cole Publishing Company: Pacific Grove, CA, USA, 2005. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 19 September 2019).
- Dormann, C.F.; Fruend, J.; Gruber, B.; Dormann, M.C. Package ‘Bipartite’. Version 2.14. Available online: https://cran.r-project.org/web/packages/lmtest/index.pdf (accessed on 20 September 2019).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H.; et al. Package ‘Vegan’. Community Ecology Package; 2013; Volume 2; pp. 1–295. [Google Scholar]
- Almeida-Neto, M.; Guimaraes, P.; Guimaraes, P.R., Jr.; Loyola, R.D.; Ulrich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Memmott, J.; Waser, N.M.; Price, M.V. Tolerance of pollination networks to species extinctions. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Burgos, E.; Ceva, H.; Perazzo, R.P.; Devoto, M.; Medan, D.; Zimmermann, M.; Delbue, A.M. Why nestedness in mutualistic networks? J. Theor. Biol. 2007, 249, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E. Detecting community structure in networks. Eur. Phys. J. B 2004, 38, 321–330. [Google Scholar] [CrossRef]
- Haenke, S.; Scheid, B.; Schaefer, M.; Tscharntke, T.; Thies, C. Increasing syrphid fly diversity and density in sown flower strips within simple vs. complex landscapes. J. Appl. Ecol. 2009, 46, 1106–1114. [Google Scholar] [CrossRef]
- Balzan, M.V.; Bocci, G.; Moonen, A.C. Augmenting flower trait diversity in wildflower strips to optimise the conservation of arthropod functional groups for multiple agroecosystem services. J. Insect Conserv. 2014, 18, 713–728. [Google Scholar] [CrossRef]
- Mkenda, P.A.; Ndakidemi, P.A.; Stevenson, P.C.; Arnold, S.E.; Belmain, S.R.; Chidege, M.; Gurr, G.M. Field margin vegetation in tropical African bean systems harbours diverse natural enemies for biological pest control in adjacent crops. Sustainability 2019, 11, 6399. [Google Scholar] [CrossRef] [Green Version]
- Zytynska, S.E.; Eicher, M.; Fahle, R.; Weisser, W.W. Effect of flower identity and diversity on reducing aphid populations via natural enemy communities. Ecol. Evol. 2021, 11, 18434–18445. [Google Scholar] [CrossRef]
- Shelton, A.M.; Badenes-Perez, F.R. Concepts and applications of trap cropping in pest management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.C.; Wang, E.; Wu, S.; Lei, Z. Application of Trap Cropping as Companion Plants for the Management of Agricultural Pests: A Review. Insects 2018, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Eubanks, M.D.; Denno, R.F. Health food versus fast food: The effects of prey quality and mobility on prey selection by a generalist predator and indirect interactions among prey species. Ecol. Entomol. 2000, 25, 140–146. [Google Scholar] [CrossRef]
- Lewinsohn, T.M.; Inácio Prado, P.; Jordano, P.; Bascompte, J.; Olesen, J.M. Structure in plant-animal interaction assemblages. Oikos 2006, 113, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, C.; Thébault, E.; Dajoz, I. Are insect pollinators more generalist than insect herbivores? Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 3027–3033. [Google Scholar] [CrossRef] [Green Version]
- Morrison, B.M.; Dirzo, R. Distinct responses of antagonistic and mutualistic networks to agricultural intensification. Ecology 2020, 101, e03116. [Google Scholar] [CrossRef]
- Pocock, M.J.; Evans, D.M.; Memmott, J. The robustness and restoration of a network of ecological networks. Science 2012, 335, 973–977. [Google Scholar] [CrossRef] [Green Version]
- Araújo, W.S.D. Global patterns in the structure and robustness of plant-herbivore networks. Front. Biogeogr. 2016, 8, e31053. [Google Scholar] [CrossRef] [Green Version]
- Robertson, C. Flowers and Insects: Lists of Visitors of Four Hundred and Fifty-Three Flowers; Science Press Printing Company: Lancaster, UK, 1929. [Google Scholar]
- Tooker, J.F.; Hauser, M.; Hanks, L.M. Floral host plants of Syrphidae and Tachinidae (Diptera) of central Illinois. Ann. Entomol. Soc. Am. 2006, 99, 96–112. [Google Scholar] [CrossRef]
- Campbell, J.W.; Kimmel, C.B.; Grodsky, S.M.; Smithers, C.; Daniels, J.C.; Ellis, J.D. Wildflower plantings harbor increased arthropod richness and abundance within agricultural areas in Florida (USA). Ecosphere 2019, 10, e02890. [Google Scholar] [CrossRef]
- Zemenick, A.T.; Kula, R.R.; Russo, L.; Tooker, J. A network approach reveals parasitoid wasps to be generalized nectar foragers. Arthropod Plant Interact. 2019, 13, 239–251. [Google Scholar] [CrossRef]
- Lu, Z.X.; Zhu, P.Y.; Gurr, G.M.; Zheng, X.S.; Read, D.M.; Heong, K.L.; Yang, Y.J.; Xu, H.X. Mechanisms for flowering plants to benefit arthropod natural enemies of insect pests: Prospects for enhanced use in agriculture. Insect Sci. 2014, 21, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Russo, L.; Debarros, N.; Yang, S.; Shea, K.; Mortensen, D. Supporting crop pollinators with floral resources: Network-based phenological matching. Ecol. Evol. 2013, 3, 3125–3140. [Google Scholar] [CrossRef] [PubMed]
- Moreira, E.F.; Boscolo, D.; Viana, B.F. Spatial heterogeneity regulates plant-pollinator networks across multiple landscape scales. PLoS ONE 2015, 10, e0123628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thébault, E.; Fontaine, C. Stability of ecological communities and the architecture of mutualistic and trophic networks. Science 2010, 329, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.E.; Rader, R. Network modularity influences plant reproduction in a mosaic tropical agroecosystem. Proc. R. Soc. B 2019, 286, 20190296. [Google Scholar] [CrossRef]
- Borchardt, K.E.; Morales, C.L.; Aizen, M.A.; Toth, A.L. Plant–Pollinator conservation from the perspective of systems-ecology. Curr. Opin. Insect Sci. 2021, 47, 154–161. [Google Scholar] [CrossRef]
- Jordán, F.; Okey, T.A.; Bauer, B.; Libralato, S. Identifying important species: Linking structure and function in ecological networks. Ecol. Modell. 2008, 216, 75–80. [Google Scholar] [CrossRef]
- Maia, K.P.; Vaughan, I.P.; Memmott, J. Plant species roles in pollination networks: An experimental approach. Oikos 2019, 128, 1446–1457. [Google Scholar] [CrossRef]
- Chakraborty, P.; Chatterjee, S.; Smith, B.M.; Basu, P. Seasonal dynamics of plant pollinator networks in agricultural landscapes: How important is connector species identity in the network? Oecologia 2021, 196, 825–837. [Google Scholar] [CrossRef]
- Crespo, A.; Aguilar, J.M.; Pintado, K.; Tinoco, B.A. Key plant species to restore plant–hummingbird pollinator communities in the southern Andes of Ecuador. Restor. Ecol. 2021, 29, e13557. [Google Scholar] [CrossRef]
- Campbell, C.; Yang, S.; Shea, K.; Albert, R. Topology of plant–pollinator networks that are vulnerable to collapse from species extinction. Phys. Rev. E 2012, 86, 021924. [Google Scholar] [CrossRef] [Green Version]
- Windsor, F.M.; Tavella, J.; Rother, D.C.; Raimundo, R.L.; Devoto, M.; Guimarães, P.R., Jr.; Evans, D.M. Identifying plant mixes for multiple ecosystem service provision in agricultural systems using ecological networks. J. Appl. Ecol. 2021, 58, 2770–2782. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laha, S.; Chatterjee, S.; Das, A.; Smith, B.; Basu, P. Selection of Non-Crop Plant Mixes Informed by Arthropod-Plant Network Analyses for Multiple Ecosystem Services Delivery Towards Ecological Intensification of Agriculture. Sustainability 2022, 14, 1903. https://doi.org/10.3390/su14031903
Laha S, Chatterjee S, Das A, Smith B, Basu P. Selection of Non-Crop Plant Mixes Informed by Arthropod-Plant Network Analyses for Multiple Ecosystem Services Delivery Towards Ecological Intensification of Agriculture. Sustainability. 2022; 14(3):1903. https://doi.org/10.3390/su14031903
Chicago/Turabian StyleLaha, Supratim, Soumik Chatterjee, Amlan Das, Barbara Smith, and Parthiba Basu. 2022. "Selection of Non-Crop Plant Mixes Informed by Arthropod-Plant Network Analyses for Multiple Ecosystem Services Delivery Towards Ecological Intensification of Agriculture" Sustainability 14, no. 3: 1903. https://doi.org/10.3390/su14031903