
Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae)


1
ECNU-Alberta Joint Laboratary for Biodiversity Study, Tiantong Forest Ecosystem National Observation and Research Station, School of Ecology and Environmental Sciences, East China Normal University, Shanghai 200241, China
2
Shanghai Institute of Pollution Control and Ecological Security, Shanghai 200092, China
*
Author to whom correspondence should be addressed.
Forests 2022, 13(6), 961; https://doi.org/10.3390/f13060961
Submission received: 27 May 2022
/
Revised: 15 June 2022
/
Accepted: 16 June 2022
/
Published: 19 June 2022
(This article belongs to the Special Issue Topicalities in Forest Ecology of Seeds)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Disordered germination is widely observed in plant species with extremely small populations (PSESPs) in China. Abies beshanzuensis M. H. Wu, a PSESP (with only three extant adults in Zhejiang Province, China) that also has poor seed germination in the field, belongs to the Pinaceae family. Pinaceae generally tend toward symbiosis with ectomycorrhizal (ECM) fungi to alleviate climate change-induced stresses. Therefore, exploring how to improve seed germination of A. beshanzuensis and whether it is an ECM species will contribute to increasing recruitment for conducting further conservation research. In this study, four temperature regimes (10/15 °C, 15/20 °C, 20/25 °C, and 25/30 °C) were selected to explore the response of seed germination to rising temperature. Based on the microscopic features of fine roots, together with molecular techniques, the mycorrhizal type of this species was identified. The seed germination of A. beshanzuensis was increased from 1–2% to around 4.5% by 14-day cold stratification and under 20/25 °C fluctuating temperature conditions. Our findings indicated that A. beshanzuensis may be endangered as a result of insufficient seedling recruitment due to poor germination under the current climate. A. beshanzuensis was confirmed as an ECM fungi-associated tree species. This study highlights the necessity of incorporating auxiliary seed treatment into population recovery programs of A. beshanzuensis, thereby better conserving the species under ongoing environmental changes.
1. Introduction
For plant species with extremely small populations (PSESPs) in China, with economic or scientific value [1,2], in addition to being subject to habitat fragmentation and excessive human destruction, climate change has disturbed the plants’ life cycles [3]. Studies have reported decreases in the reproductive capacity of numerous PSESPs [4]. Of the first batch of 120 PSESPs species listed, only 28 have been recorded with usable germination information [2], and little is known about the consequences of global warming on the seed germination of PSESPs [5]. Therefore, acquiring knowledge on seed germination under global warming is an urgent need for further restoration efforts of PSESPs.
Seed germination is the process in which a quiescent embryo activates and grows into an autotrophic seedling [6]. As a vital stage of plant life history, seed germination can determine subsequent population dynamics [7]. However, this process is considered a bottleneck, exhibits high vulnerability [8], and is subject to an unpredictable external environment (including factors such as temperature, humidity, and light) [9,10]. Among these factors, temperature is a critical driver affecting biochemical reactions in seeds when water is in plentiful supply [11,12]. The suitable temperature for seed germination is driven by the ecological conditions of the parental plants and the seed maturation environment [13], and affects the germination time and germination rate [14]. The germination percentage and germination time, two crucial traits of seed germination, determine the number of seeds that sprout into seedlings and when seedlings begin to emerge in the environment [14]. Seed mass, as a plant phenotypic characteristic, is also related to germination and post-germination performance [15]. Studies have shown that plants growing in hostile environments tend to produce larger seeds [7], although producing heavier seeds is energetically costly and results in a lower seed yield [16]. This is because heavier seeds exhibit enhanced successful emergence from litter layers and produce larger seedlings with higher stress resistance [17]. Environmental conditions drive plants to make tradeoffs between seed mass (fecundity) and quantity (stress resistance) [18,19]. These tradeoffs allow seeds to tolerate stresses and germinate under suitable conditions, thereby ensuring population recruitment.
Abies beshanzuensis M. H. Wu (Pinaceae) was officially identified as a new species in 1976 and then listed as one of the PSESPs needing priority protection [20]. The discovery of this species disproves the supposition that there are no firs in the subtropical low–mid-mountain area of China, which is of far-reaching significance for obtaining a better understanding of the paleoclimate, paleogeography, and the evolution of flora in this region [21]. Although the distribution of Abies once extended to southern China before the Quaternary glaciation [22], significant global warming has pushed the population to the upper part of Baishanzu Mountain [23]. Today, only three adult individuals of A. beshanzuensis are extant in the subtropical forest (Zhejiang province; 27°4′–27°5′ N, 119°3′–119°6′ E; 1750 m) [24]. Global warming poses an enormous threat to the growth and habitat of this species [22]. A large number of previous studies were conducted with a focus on restoring the population of A. beshanzuensis through vegetative reproduction, such as through grafting and tissue culture methods [25]. However, to maintain the genetic diversity of plant species, seed propagation is a more effective method [4]. The staff of the Baishanzu Management Office carried out seed germination experiments on A. beshanzuensis in the field. The highest germination percentage was only 1.48% [26]. According to other studies on Abies, the seed germination performance of A. beshanzuensis may be improved by providing more favorable ambient conditions. For example, the germination percentage of Abies yuanbaoshanensis Y. J. Lu et L. K. Fu in an incubator was 18.9%, which was significantly higher than that in open air (6.8%) [27]. Another experiment showed that alternating temperature regimes could further increase seed germination in Abies chensiensis Tiegh. [28]. Therefore, exploring the effects of different temperature cycles on seed germination, in order to elucidate the responses of seeds to global warming and population dynamics, is of great significance.
Merely improving the seed germination of A. beshanzuensis may not be enough to guarantee its population recruitment. Recently, accumulating studies have highlighted the previously neglected role of ectomycorrhizal (ECM) fungi in alleviating climate change-induced stresses via enhancing host plant tolerance [29,30], especially at the vulnerable seedling stage. A symbiotic relationship (mutualism) is likely to form when seeds germinate into seedlings and encounter compatible ECM fungi from the surrounding environment [31]. However, it should be noted that ECM fungi usually have high host specificity, whereby ECM fungi only benefit the direct descendants in the immediate vicinity of their parent tree species [32]. Different ECM fungi have different effects and can improve plant adaptation by cooperating with other biotic and abiotic factors [25]. Pinaceae generally tend to interact with ECM fungi under certain conditions [33,34]. Whether A. beshanzuensis, an original member of Pinaceae [21], is an ECM plant should be further confirmed based on microscopic characteristics and molecular identification of fine roots. If this is the case, in combination with increased seed germination, ECM facilitation should be incorporated into restoration programs to better increase the population recruitment of this critically endangered tree species.
In the present study, we investigated the contribution of auxiliary seed treatment to improving A. beshanzuensis seed germination (a critical phase) under different temperature regimes, and identified ECM fungi colonizing the roots of this species. Specifically, our aim was to determine the following: (1) Compared with other Abies endemic to China, what is the germination performance of A. beshanzuensis? (2) What is the effect of different temperature regimes on seed germination in A. beshanzuensis? (3) Is A. beshanzuensis an ECM tree? These basic data will have practical implications for the population recovery of A. beshanzuensis.
2. Materials and Methods
2.1. Study Species and Sample Collection
The grafted A. beshanzuensis was selected as the focal tree species (Zhejiang Province; 27°4′–27°5′ N, 119°3′–119°6′ E; 1590 m). The limited seeds collected from native trees in 2019 were not sufficient for the experiment, and an analysis determined that the grafted A. beshanzuensis had a high degree of genetic consistency with maternal plants [21]. The wild germination of A. beshanzuensis generally occurs in late spring [20], when the temperature fluctuation is obvious (Figure 1, data were obtained from (http://www.cdc.noaa.gov, accessed on 5 October 2020). Abies beshanzuensis var. ziyuanensis (L. K. Fu et S. L. Mo) L. K. Fu et Nan Li, like A. beshanzuensis, is a “living plant fossil” left over from the Quaternary glaciation [22]. There are more than 1000 A. ziyuanensis trees in Hunan Province (26°1′–26°3′ N, 114°1′–114°4′ E; 1430 m) [23]. A. ziyuanensis are distributed at approximately similar altitudes and latitudes to A. beshanzuensis, but are less endangered than A. beshanzuensis [25]. Therefore, after comprehensive consideration, A. ziyuanensis was selected as a reference to understand the germination performance of A. beshanzuensis.
We collected seeds from three grafted A. beshanzuensis and five A. ziyuanensis. Both types of seeds were collected as they began to fall spontaneously in late October 2019. We also randomly selected five A. beshanzuensis trees from three altitudes (951, 1400, and 1590 m) that were artificially cultivated in 1992 in order to collect their fine roots. Fine roots were carefully collected after removing the weeds and litter around the target tree. The root samples were taken back to the laboratory in an icebox and kept at 4 °C for further analyses.
2.2. Thousand-Grain Weight
Thousand-grain weight refers to the embryo and endosperm plus the seed coat (excluding the seed wing) in the natural dry state. Eight groups of 100 pure seeds were selected stochastically from the samples for measurement. When the coefficient of variance (CV) exceeded 4%, the aforementioned process was repeated to obtain the true weight value [28].
This study also collected seed weights and optimal germination percentages of other Abies endemic to China from the literature, including Abies georgei Orr, Abies georgei Orr var. smithii (Viguie et Gaussen) Cheng et L., Abies fargesii Franch., A. chensiensis, A. yuanbaoshanensis, and Abies holophylla Maxim., to further understand the status of A. beshanzuensis. Among them, A. beshanzuensis, A. ziyuanensis, and A. yuanbaoshanensis were listed as PSESPs.
2.3. Seed Viability Test
The tetrazolium test was repeated three times with a group of 50 seeds from two populations (A. beshanzuensis and A. ziyuanensis). Seeds were cut longitudinally and half were left to soak in 1% TTC solution (2, 3, 5-triphenyl tetrazolium chloride) at 30 °C for staining 12 h after immersion in ultrapure water for 18 h. The seed viability was evaluated based on the number of uniformly stained seeds observed.
2.4. Seed Germination Experiment
Sterilized seeds were layered with moistened sand in three plastic boxes with holes drilled into them for drainage. The plastic boxes were stored in a refrigerator at a constant temperature (4 °C) to enable cold stratification to break seed physiological dormancy. The seed conditions and humidity were checked every week. The stratification times were 28, 14, and 0 days (dry storage under cold conditions).
In April 2020, we soaked seeds that had finished the stratification treatment in 30 °C water for 24 h and sterilized their surfaces with 1% hydrogen peroxide solution for 1 h. Seeds were germinated with a humidity of 75% and a light source (8 h of light, 16 h of darkness) in four incubators set to 10/15 °C, 15/20 °C, 20/25 °C, and 25/30 °C (8 h of high temperature, 16 h of low temperature). For each temperature, four replicates of 50 seeds (4800 seeds = 50 seeds × 4 replicates × 12 treatments × 2 species) were cultivated evenly in 90 mm diameter Petri dishes on two layers of filter paper. We kept the filter paper moist and randomly placed Petri dishes in the incubators. We also cleaned the moldy seeds promptly and replaced the filter paper every five days. Monitoring had been carried out since the first day of germination. The criterion for successful germination was that the radical length reached half the seed length. The experiment was terminated when no germination had occurred for 7 successive days [35]. The final germination percentage (total germinated seed number divided by the total number of seeds tested × 100) and germination timing were recorded.
2.5. Fine Root Identification and Molecular Analysis
The fine root samples of A. beshanzuensis were gently rinsed with running water to remove adhered soil. The root tips were examined under a microscope to determine whether they were infected with ECM fungi [36].
We further investigated the ECM fungal community present on roots with molecular techniques. Over 500 mg of root tips were surface-sterilized and homogenized in liquid N. We extracted DNA using a plant DNA kit. The obtained genome could be directly used for PCR. The PCR reaction was performed with the universal fungal primers ITS3 (GCATCGATGAAGAACGCAGC) and ITS4 (TCCTCCGCTTATTGATATGC). The PCR mixture was recovered, and the target DNA fragments were eluted with TE buffer. After obtaining the original data, the sequencing quality was evaluated and controlled. The USEARCH process and the Unite database were merged to cluster sequences into operational taxonomic units (OTUs) with a similarity threshold of 97% [37]. The SUB.SAMPLE command in mothur was used to obtain a normalized OTU table for analysis.
2.6. Statistical Analyses
Analysis of variance (ANOVA) was used to test whether there were significant differences between seed germination (A. beshanzuensis and A. ziyuanensis) rates under different temperatures (10/15 °C, 15/20 °C, 20/25 °C, and 25/30 °C), and Tukey’s test was used to further determine in which of the two groups of treatments such differences existed. Prior to analyzing the data, the final germination percentage (Y) and the initial germination time were transformed using sqrt. The aforementioned statistical analyses were performed in R.
The endophytic fungal OTUs of root tissues were taxonomically parsed based on the FUNGuild database (http://www.stbates.org/guilds/app.php, accessed on 16 March 2021) and categorized into trophic guilds [38].
3. Results
3.1. Seed Mass and Viability
The thousand-grain weight of A. beshanzuensis was 33.76 g (CV = 0.058% < 4.000%), and the thousand-grain weight of A. ziyuanensis was 33.70 g (CV = 0.061% < 4.000%). The population studies of Abies endemic to China showed a weak negative relationship between the germination capacity and seed mass (p = 0.128, R2 = 0.232). As shown in Figure 2, A. beshanzuensis had an extremely low germination rate.
The tetrazolium experiment results showed that the seed viability of A. beshanzuensis was 4.67 ± 0.66%, and that of A. ziyuanensis was 10.00 ± 1.15%. It was worth noting that A. ziyuanensis had the same thousand-grain weight as A. beshanzuensis, but the seed viability and the optimum seed germination percentage of A. ziyuanensis were twice those of A. beshanzuensis (Figure 3). This figure also shows that proper treatment can promote the germination of viable seeds.
3.2. Seed Germination Percentage
The initial germination time advanced as the incubator temperature increased (Figure 4a). The optimal final germination percentage of A. beshanzuensis was obtained under the fluctuating temperature conditions of 20/25 °C (with 14-day cold stratification) (Figure 4a). The optimal germination condition for A. ziyuanensis was to cultivate the seeds at 15/20 °C (with 28-day cold stratification) (Figure 4b). Compared with A. ziyuanensis, the germination of A. beshanzuensis continued to advance with the increase in the temperature (Figure 4).
3.3. ECM Fungi Present on Fine Roots of A. beshanzuensis
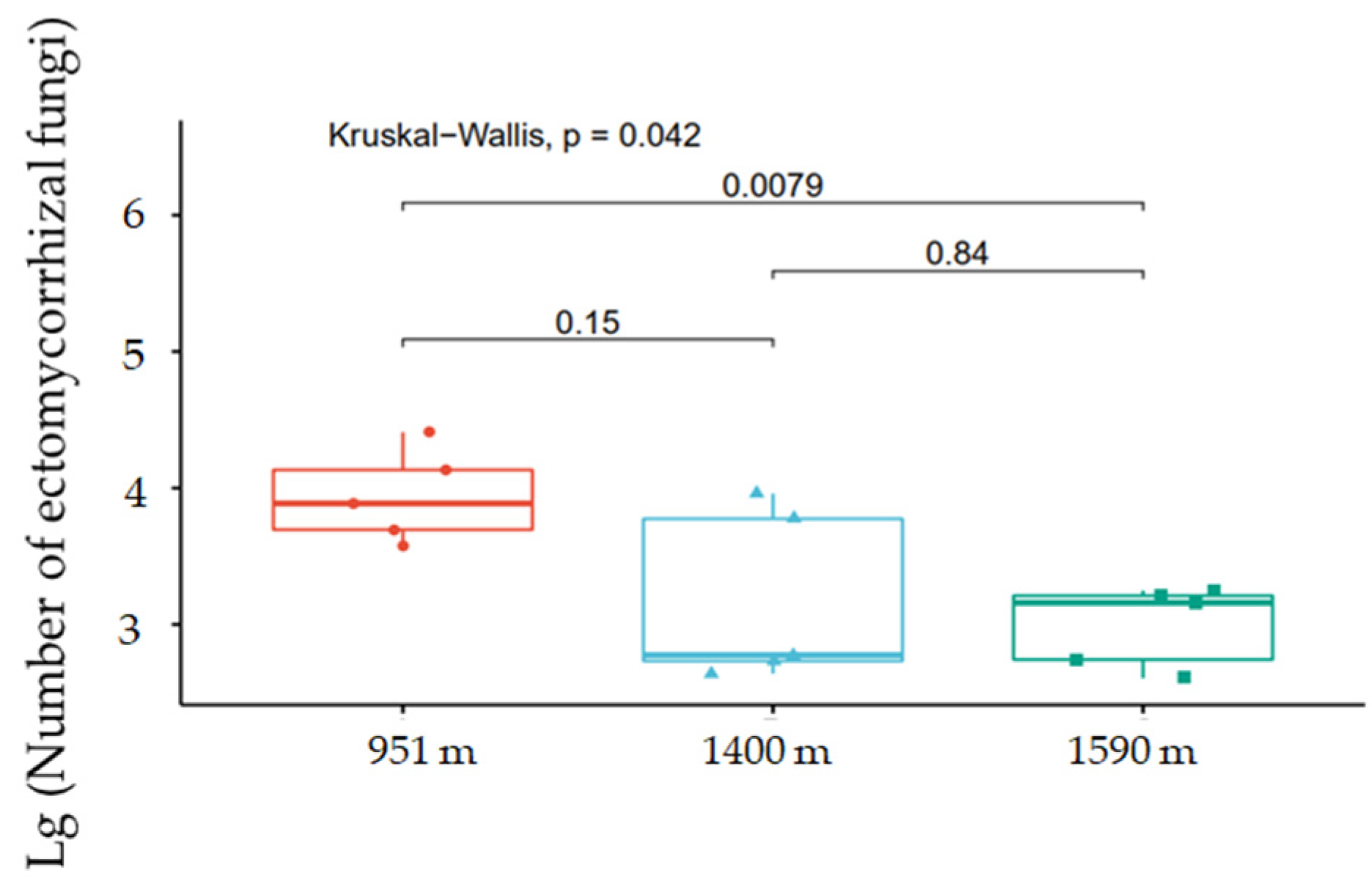
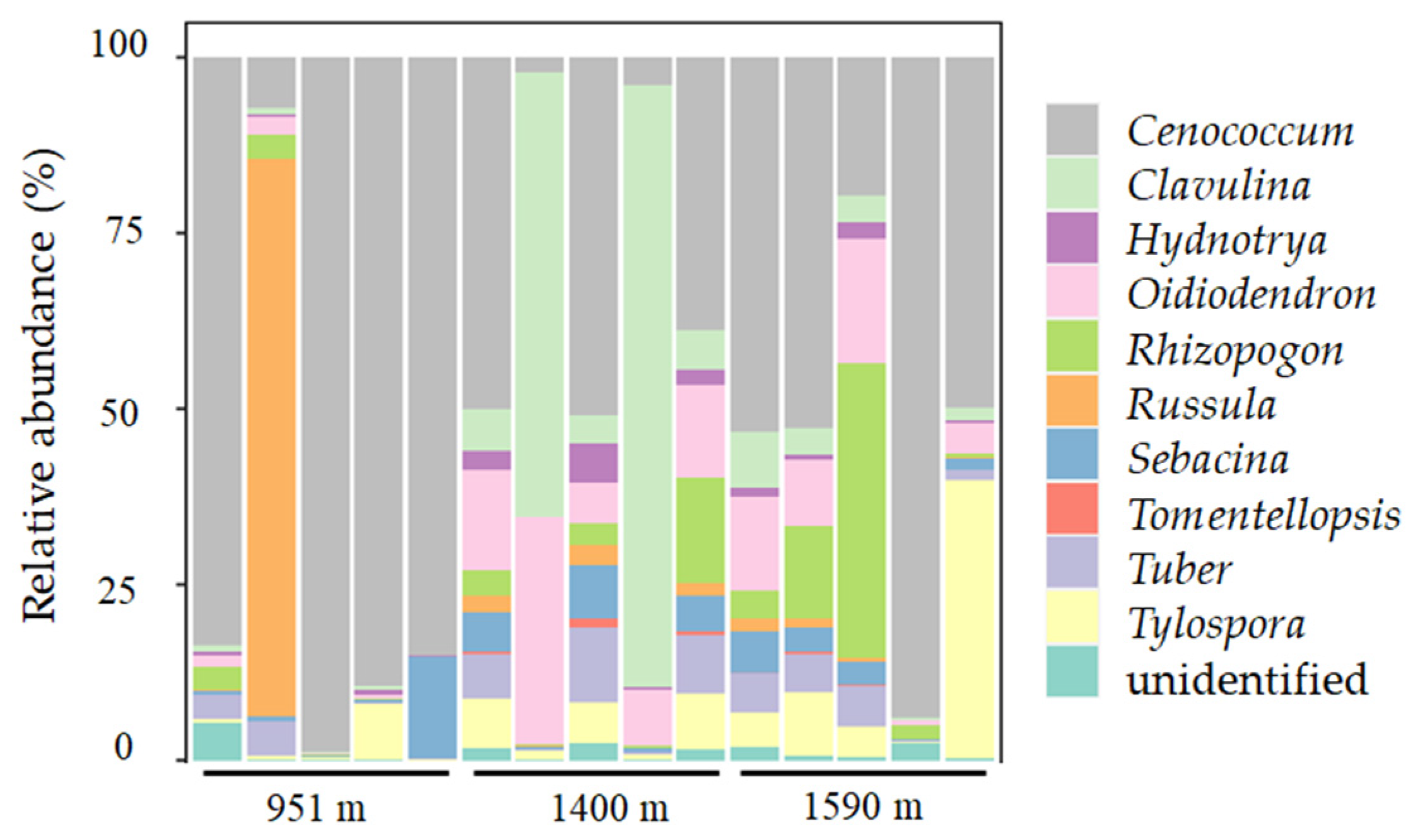
A. beshanzuensis was determined to be an ECM plant by observing its microscopic features (Figure 5). Short roots that grow slowly allow ECM fungi to form an association. Hyphae reached the root surface and formed a firm sheath called the mantle in order to resist pathogen stresses, while penetrating the cortex to form a Hartig net to obtain water and nutrients. The root tips of A. beshanzuensis appeared brown and black (Figure 5). The Kruskal–Wallis rank sum test was used to explore the log-transformed differences in the number of ECM fungi in the roots of A. beshanzuensis at different altitudes, and it was found that there was a significant difference between plants at 951 m and 1590 m (Figure 6). However, the number of ECM fungi in the fine root tissues of A. beshanzuensis was relatively high at high and low altitudes. At least 10 ECM fungal species (belonging to families in the Ascomycota and Basidiomycota) inhabited the roots and formed symbiotic combinations (Figure A2).
4. Discussion
4.1. Relationships between Seed Mass and Germination Percentage
Three species of Abies that were listed as PSESPs (AB: A. beshanzuensis, AZ: A. ziyuanensis, and AY: A. yuanbaoshanensis) tended to have relatively low seed germination no matter the seed size (Figure 2), implying that these species were not capable of at least maintaining population regeneration by adopting different seed strategies. Under unfavorable conditions, seed germination and seedling survival are closely related to the nutrient reserves in seeds, which increase with seed weight [19]. A. beshanzuensis, which prefers growing in a humid and cool environment, produces heavier seeds to ensure germination and adaptability under the background of global warming (Figure 2), indicating that this species is attempting to rescue itself under severe stress. Unfortunately, this strategy does not seem to be successful at present. Compared with A. ziyuanensis, the germination percentage of A. beshanzuensis was, indeed, poor (Figure 2). The results of the seed viability test also indirectly confirmed this point (Figure 3). This may be because the minimum viable A. beshanzuensis population for long-term survival in the wild is much lower than that of A. ziyuanensis [22]. The shrinking population size that accompanies genetic simplification could weaken the ability of plants to produce sufficient quantities of pollen, resulting in poor germination performance in A. beshanzuensis [21]. This is the greatest challenge that A. beshanzuensis faces. This finding underscores the urgent need for artificial measures to protect this endangered species.
4.2. Effects of Warming on Seed Germination in A. beshanzuensis
The optimum germination temperature and stratification time of A. beshanzuensis were different from those of A. ziyuanensis (Figure 3 and Figure 4). The optimal temperature for seed germination of A. beshanzuensis was 20–25 °C (Figure 3). However, it is difficult for the temperature of its habitat to stabilize at 20–25 °C from March to April (Figure 1). In addition, the dormancy of A. beshanzuensis could be broken after 14-day cold stratification, indicating that the dormancy degree of A. beshanzuensis was lower than that of A. ziyuanensis (Figure 3 and Figure 4). A temporary suitable temperature is likely to induce the germination of A. beshanzuensis, forcing new seedlings to be exposed to adverse environmental conditions earlier. Therefore, compared with the seed germination of A. ziyuanensis, the seed germination of A. beshanzuensis will face more challenges under the backdrop of global warming.
Mild increases in temperature promoted the final germination percentage and advanced the initial germination time of A. beshanzuensis (Figure 3, Figure 4 and Figure A1). Compared with A. ziyuanensis, the germination of A. beshanzuensis continued to advance with the rise in temperature (Figure 4), which would affect the growth of seedlings. This was because the seed germination of many alpine plants is induced by the gradual increase in temperature as snow melts [39]. This process is completely irreversible, and once germination has started, the seed must confront the risks it encounters [7]. In the field, the point in the year when temperatures are favorable for germination is advanced with warming, advancing the time of emergence, despite the possibility of early spring frosts (Figure 1). When seeds begin to germinate, subsequent temperature fluctuations make it difficult for seeds to smoothly progress through the process from seed germination to seedling establishment in field environments [39]. Therefore, the positive effect of mild increases in temperature on the regeneration of A. beshanzuensis seedlings (increase in germination rate) was smaller than the negative effect (an increase in the possibility of early spring frost stress due to an earlier initial germination time). Having a large number of seedlings is a prerequisite for carrying out subsequent protection measures. Therefore, it is necessary to utilize auxiliary seed treatments, such as artificial incubators, to create a suitable environment to successfully cultivate a large number of seedlings.
4.3. ECM Root-Tip Communities
A. beshanzuenesis was identified as an ECM species based on microscopic characteristics and the molecular identification of fine roots (Figure 5 and Figure A2). The role of ECM symbioses has key implications for slowing the progression of endangered species extinction [36,40]. Specifically, ECM fungi present on root tips of A. beshanzuenesis could facilitate plant growth through robust hyphal networks that could acquire nutrients in resource-poor conditions, as well as create a dense mantle that surrounds fine roots (Figure 5) to increase stress resistance [38,41]. This is likely why A. beshanzuensis, which prefers growing in humid and cool environments, has a greater dependence on fungi at lower altitudes with higher temperatures (Figure 6). In addition, the ECM fungi that were colonized in the roots of A. beshanzuenesis were categorized into 2 phyla and 10 genera via molecular techniques (Figure A2). Colonized ECM fungal species had different functions and lifestyles [42]. Clavulina is widely distributed at high elevations [43]. Russula appears to respond negatively to warming [44]. Cenococcum penetrating into root tissue can form a dense mantle that protects the host against stresses [45]. These ECM fungi enter the cortex cells of root tissue and result in marked enhancements in water and nutrient uptake, drought resistance, and seedling biomass at competitive sites [46,47]. Therefore, A. beshanzuensis may assemble the ectomycorrhizal fungal network to improve its adaptability to the environment (Figure A2). The results of this study have provided hints that the complementary benefits of ECM fungi could be amplified to aid A. beshanzuenesis seedling establishment. In future research, ECM fungi will be selected and cultivated from the root tissues of parental plants in order to prepare fungal inoculum and inoculate A. beshanzuensis seedlings living in the nursery. This will lay a foundation for the application of field ex situ conservation experiments.
5. Conclusions
The smooth passage of the process from seed germination to seedling establishment is critical to ensure subsequent plant development. However, A. beshanzuensis, a critically endangered species, has constraints on natural regeneration. Therefore, auxiliary seed treatment of A. beshanzuensis must be considered in order to protect this endangered species. The experiments performed in this study have shown that the seed germination rate of A. beshanzuensis can be increased from 1–2% to around 4.5% by 14-day cold stratification and a 20/25 °C temperature regime. In addition, A. beshanzuensis has been identified as an ECM tree. ECM symbiosis provides strong support for both partners in natural conditions, and this relationship should be exploited to further restoration efforts. In the present study, suitable ECM fungi were identified in the root tips of parental trees. Moving forward, we will integrate molecular tools with physiological experiments to select appropriate ECM fungi and then inoculate them into seedlings. In practice, this study proposes that the improved recovery of endangered species may be achieved through multiple techniques, thereby shedding more light on population dynamics.
Author Contributions
Y.L. (Yu Liu) and R.H. conceived and designed the experiments; R.H. performed the experiment and data analysis; R.H., H.X. and J.Z. contributed to manuscript preparation; R.H. and Y.L. (Yu Liu) wrote the original draft, performed the analysis, and supplied the discussion; and R.H., Y.L. (Yajing Liu) and S.J. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31670531 and 32071645). Additional funding was provided by the Fengyangshan–Baishanzu National Nature Reserve (2020FWR0208).
Data Availability Statement
The data that support the findings of this study are available from the author upon reasonable request.
Acknowledgments
We thank Yougui Wu and Zhicheng Zhu for their assistance in the field and for their suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Cumulative germination percentages under different cold stratification treatment: (a) Abies beshanzuensis (21-day cold stratification); (b) Abies beshanzuensis (28-day cold stratification); (c) Abies ziyuanensis (14-day cold stratification); (d) Abies ziyuanensis (21-day cold stratification).
Figure A1.
Cumulative germination percentages under different cold stratification treatment: (a) Abies beshanzuensis (21-day cold stratification); (b) Abies beshanzuensis (28-day cold stratification); (c) Abies ziyuanensis (14-day cold stratification); (d) Abies ziyuanensis (21-day cold stratification).

Figure A2.
Relative abundances of ectomycorrhizal fungi species (genus level) on resident Abies beshanzuensis tree roots as detected by molecular analysis.
Figure A2.
Relative abundances of ectomycorrhizal fungi species (genus level) on resident Abies beshanzuensis tree roots as detected by molecular analysis.

References
- Volis, S. How to conserve threatened Chinese plant species with extremely small populations? Plant Divers. 2016, 38, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Wade, E.M.; Nadarajan, J.; Yang, X.; Ballesteros, D.; Sun, W.; Pritchard, H.W. Plant species with extremely small populations (PSESP) in China: A seed and spore biology perspective. Plant Divers. 2016, 38, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Cai, L.; Liu, D.; Chen, G.; Gratzfeld, J.; Sun, W. China’s conservation program on Plant Species with Extremely Small Populations (PSESP): Progress and perspectives. Biol. Conserv. 2020, 244, 108535. [Google Scholar] [CrossRef]
- Sha, D.; Yanni, W.; Kunlin, W.; Lin, F.; Lin, L.; Songjun, Z. Breeding characteristics and artificial propagation of 14 species of Wild Plant with Extremely Small Populations (WPESP) in China. Biodivers. Sci. 2020, 28, 385–400. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, G.; Grumbine, R.E.; Dao, Z.; Sun, W.; Guo, H. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Donohue, K.; De Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, Postgermination Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: London, UK, 2014. [Google Scholar]
- Koutecká, E.; Lepš, J. Effect of Light and Moisture Conditions and Seed Age on Germination of Three Closely Related Myosotis Species. Folia Geobot. 2009, 44, 109–130. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. (Eds.) Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA; Berlin/Heidelberg, Germany; Dordrecht, The Netherlands; London, UK, 2013; p. 392. [Google Scholar]
- Cochrane, A.; Hoyle, G.L.; Yates, C.J.; Wood, J.; Nicotra, A.B. Predicting the impact of increasing temperatures on seed germination among populations of Western Australian Banksia (Proteaceae). Seed Sci. Res. 2014, 24, 195–205. [Google Scholar] [CrossRef]
- Orrù, M.; Mattana, E.; Pritchard, H.W.; Bacchetta, G. Thermal thresholds as predictors of seed dormancy release and germination timing: Altitude-related risks from climate warming for the wild grapevine Vitis vinifera subsp. sylvestris. Ann. Bot. 2012, 110, 1651–1660. [Google Scholar] [CrossRef] [Green Version]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. Camb. Philos. Soc. 2015, 90, 31–59. [Google Scholar] [CrossRef]
- Liu, Y.; El-Kassaby, Y.A. Timing of seed germination correlated with temperature-based environmental conditions during seed development in conifers. Seed Sci. Res. 2014, 25, 29–45. [Google Scholar] [CrossRef]
- Eriksson, O. Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis. Acta Oecologica 1999, 20, 61–66. [Google Scholar] [CrossRef]
- Kidson, R.; Westoby, M. Seed mass and seedling dimensions in relation to seedling establishment. Oecologia 2000, 125, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.C.; Noland, T.L.; Morneault, A.E. The Effects of Seed Mass on Germination, Seedling Emergence, and Early Seedling Growth of Eastern White Pine (Pinus strobus L.). New For. 2006, 32, 33–49. [Google Scholar] [CrossRef]
- Bu, H.; Chen, X.; Xu, X.; Liu, K.; Jia, P.; Du, G. Seed mass and germination in an alpine meadow on the eastern Tsinghai–Tibet plateau. Plant Ecol. 2007, 191, 127–149. [Google Scholar] [CrossRef]
- Hantsch, L.; Bruelheide, H.; Erfmeier, A. High phenotypic variation of seed traits, germination characteristics and genetic diversity of an invasive annual weed. Seed Sci. Res. 2013, 23, 27–40. [Google Scholar] [CrossRef]
- Liu, X.T. Predicting the Suitable Distribution Area of Abies beshanzuensis: To Explor the Potential Dispersion of the Plant Species with Extremely Small Populations. Master’s Thesis, Zhejiang Normal University, Jinhua, China, 2019. [Google Scholar]
- Ai, J.G. Studies on the Endangerment Mechanism and Conservation Strategies for Abies beshanzuensis. Master’s Thesis, Zhejiang University, Hangzhou, China, 2005. [Google Scholar]
- Li, X.-X.; Tao, C.; Wang, Q.-C.; Cui, G.-F. Characteristics of geographic distribution of four critically endangered species of Abies in subtropical China and its relationship with climate. Chin. J. Plant Ecol. 2012, 36, 1154–1164. [Google Scholar] [CrossRef]
- Liu, R.; Wang, C.J.; He, J.; Zhang, Z.X. Analysis of geographical distribution of Abies in China under climate change. Bull. Bot. Res. 2018, 38, 37–46. [Google Scholar]
- He, F. Price of prosperity: Economic development and biological conservation in China. J. Appl. Ecol. 2009, 46, 511–515. [Google Scholar] [CrossRef]
- Rao, L.B. Study on Conservation Genetics of Five Endangered Abies Species by Marker Technology of Biochip. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2009. [Google Scholar]
- Wu, Y.G.; Rao, L.B.; Chen, D.L.; Ye, Z.L. Artificial seedling raising of Abies beshanzuensis seed. J. Anhui Agric. Sci. 2010, 38, 12038–12039. [Google Scholar] [CrossRef]
- Tang, R.Q.; Li, X.K.; Ou, Z.L.; Li, R.T. The fruiting characteristics and reproductive capacity of seeds of Abies yuanbaoshanenis. Bull. Bot. Res. 2001, 21, 404–408. [Google Scholar]
- Lai, J.S.; Li, Q.M.; Xie, Z.Q. Seed germination characteristic of the endangered plant Abies chensiensis. Acta Phytoecol. Sin. 2003, 27, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Classen, A.T.; Sundqvist, M.K.; Henning, J.A.; Newman, G.S.; Moore, J.A.M.; Cregger, M.A.; Moorhead, L.C.; Patterson, C.M. Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 2015, 6, 1–21. [Google Scholar] [CrossRef]
- Dalong, M.; Luhe, W.; Guoting, Y.; LiQiang, M.; Chun, L. Growth response of Pinus densiflora seedlings inoculated with three indigenous ectomycorrhizal fungi in combination. Braz. J. Microbiol. 2011, 42, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Correia, M.; Heleno, R.; Vargas, P.; Rodríguez-Echeverría, S. Should I stay or should I go? Mycorrhizal plants are more likely to invest in long-distance seed dispersal than non-mycorrhizal plants. Ecol. Lett. 2018, 21, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Hayward, J.; Horton, T.R.; Pauchard, A.; Nuñez, M.A. A single ectomycorrhizal fungal species can enable a Pinus invasion. Ecology 2015, 96, 1438–1444. [Google Scholar] [CrossRef]
- Murata, M.; Kanetani, S.; Nara, K. Ectomycorrhizal fungal communities in endangered Pinus amamiana forests. PLoS ONE 2017, 12, e0189957. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Wang, S.; Wei, X.; Jiang, M. Sensitivity of seed germination to temperature of a relict tree species from different origins along latitudinal and altitudinal gradients: Implications for response to climate change. Trees 2019, 33, 1435–1445. [Google Scholar] [CrossRef]
- Teste, F.P.; Karst, J.; Jones, M.D.; Simard, S.W.; Durall, D.M. Methods to control ectomycorrhizal colonization: Effectiveness of chemical and physical barriers. Mycorrhiza 2006, 17, 51–65. [Google Scholar] [CrossRef]
- Veach, A.M.; Yip, D.; Engle, N.L.; Yang, Z.K.; Bible, A.; Morrell-Falvey, J.; Tschaplinski, T.J.; Kalluri, U.C.; Schadt, C.W. Modification of plant cell wall chemistry impacts metabolome and microbiome composition in Populus PdKOR1 RNAi plants. Plant Soil 2018, 429, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Bernareggi, G.; Carbognani, M.; Mondoni, A.; Petraglia, A. Seed dormancy and germination changes of snowbed species under climate warming: The role of pre- and post-dispersal temperatures. Ann. Bot. 2016, 118, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2010, 17, 2145–2161. [Google Scholar] [CrossRef]
- Baird, R.; Stokes, C.E.; Frampton, J.; Smith, B.; Watson, C.; Pilgrim, C.; Scruggs, M. Diversity and density of the EM fungal community present in high elevation Fraser fir forests of Great Smoky Mountains National Park. N. Am. Fungi 2014, 9, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Teste, F.P.; Simard, S.W.; Durall, D.M.; Guy, R.D.; Jones, M.; Schoonmaker, A.L. Access to mycorrhizal networks and roots of trees: Importance for seedling survival and resource transfer. Ecology 2009, 90, 2808–2822. [Google Scholar] [CrossRef]
- Veach, A.M.; Stokes, C.E.; Knoepp, J.; Jumpponen, A.; Baird, R. Fungal communities and functional guilds shift along an elevational gradient in the southern appalachian mountains. Microbial Ecology 2017, 76, 156–168. [Google Scholar] [CrossRef]
- Fernandez, C.; Nguyen, N.H.; Stefanski, A.; Han, Y.; Hobbie, S.E.; Montgomery, R.A.; Reich, P.; Kennedy, P.G. Ectomycorrhizal fungal response to warming is linked to poor host performance at the boreal-temperate ecotone. Glob. Chang. Biol. 2016, 23, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; May, T.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2009, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Pec, G.J.; Simard, S.W.; Cahill, J.F.; Karst, J. The effects of ectomycorrhizal fungal networks on seedling establishment are contingent on species and severity of overstorey mortality. Mycorrhiza 2020, 30, 173–183. [Google Scholar] [CrossRef]
Figure 1.
Temperature from March to April in 2020 at the nearest station to Baishanzu Mountain.

Figure 2.
Compared with other Abies endemic to China, Abies beshanzuensis had low germinability. AB: Abies beshanzuensis, AC: Abies chensiensis, AF: Abies fargesii, AG: Abies georgei, Ag: Abies georgei var. smithii, AH: Abies holophylla Maxim, AY: Abies yuanbaoshanensis, AZ: Abies ziyuanensis. The blue-colored area is the 95% confidence interval.
Figure 2.
Compared with other Abies endemic to China, Abies beshanzuensis had low germinability. AB: Abies beshanzuensis, AC: Abies chensiensis, AF: Abies fargesii, AG: Abies georgei, Ag: Abies georgei var. smithii, AH: Abies holophylla Maxim, AY: Abies yuanbaoshanensis, AZ: Abies ziyuanensis. The blue-colored area is the 95% confidence interval.

Figure 3.
Variation in the final germination percentage of Abies beshanzuensis (a) and Abies ziyuanensis (b) under different treatments (mean ± SE). Red solid lines represent seed vigor as measured by a tetrazolium test. The bar is not visible for treatments with no seed germination.
Figure 3.
Variation in the final germination percentage of Abies beshanzuensis (a) and Abies ziyuanensis (b) under different treatments (mean ± SE). Red solid lines represent seed vigor as measured by a tetrazolium test. The bar is not visible for treatments with no seed germination.

Figure 4.
Cumulative germination percentages under optimal treatment: (a) Abies beshanzuensis (14-day cold stratification) and (b) Abies ziyuanensis (28-day cold stratification).
Figure 4.
Cumulative germination percentages under optimal treatment: (a) Abies beshanzuensis (14-day cold stratification) and (b) Abies ziyuanensis (28-day cold stratification).

Figure 5.
Two typical ectomycorrhizal structures of the fine roots of Abies beshanzuensis. (a) Short mycorrhizal lateral roots, which is called heterorhizy, supported by a network of thicker, long roots for resisting pathogen stresses. (b) Fungal hyphae enter the cortex to form a Hartig network, while forming a firm sheath.
Figure 5.
Two typical ectomycorrhizal structures of the fine roots of Abies beshanzuensis. (a) Short mycorrhizal lateral roots, which is called heterorhizy, supported by a network of thicker, long roots for resisting pathogen stresses. (b) Fungal hyphae enter the cortex to form a Hartig network, while forming a firm sheath.

Figure 6.
Differences in the number of ectomycorrhizal fungi in Abies beshanzuensis at different altitudes.
Figure 6.
Differences in the number of ectomycorrhizal fungi in Abies beshanzuensis at different altitudes.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, R.; Liu, Y.; Zhang, J.; Xing, H.; Jiang, S.; Liu, Y. Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae). Forests 2022, 13, 961. https://doi.org/10.3390/f13060961
AMA Style
Hu R, Liu Y, Zhang J, Xing H, Jiang S, Liu Y. Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae). Forests. 2022; 13(6):961. https://doi.org/10.3390/f13060961
Chicago/Turabian StyleHu, Rui, Yajing Liu, Jiaxin Zhang, Hua Xing, Sha Jiang, and Yu Liu. 2022. "Auxiliary Seed Treatment Is Necessary to Increase Recruitment of a Critically Endangered Species, Abies beshanzuensis (Pinaceae)" Forests 13, no. 6: 961. https://doi.org/10.3390/f13060961
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.