
Genetic, Morphological, and Environmental Differentiation of an Arid-Adapted Oak with a Disjunct Distribution

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and Microsatellite Protocol
2.3. Statistical Analysis of Genetic Data
2.4. Morphometric Methods
2.5. Environmental Differentiation and Influence on Genetic Variation
2.6. Estimation of Divergence between East and West Regions Using ABC and Genetic Data
3. Results
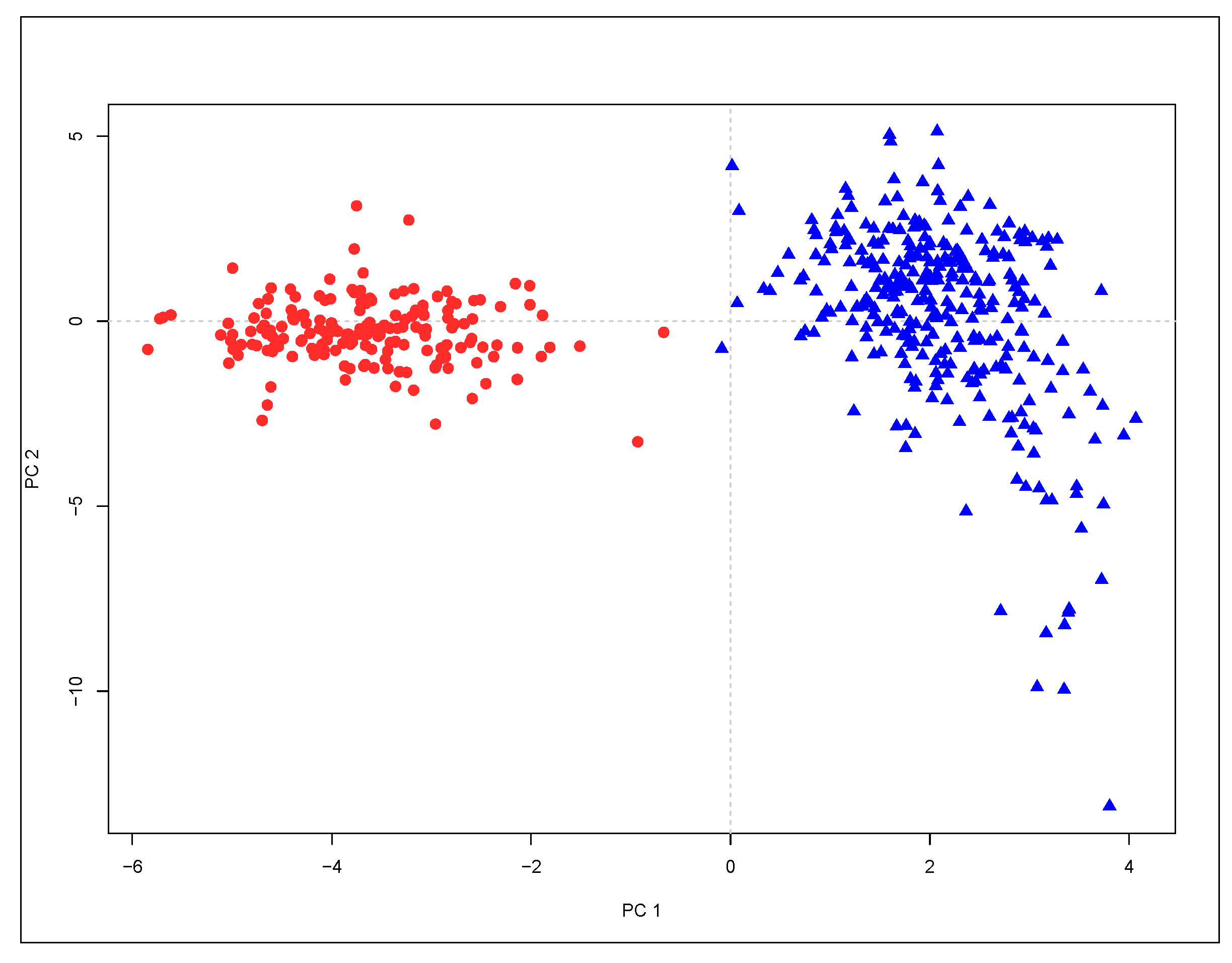
3.1. Genetic Variation and Differentiation
3.2. Morphological Variation
3.3. Environmental Differentiation, Variation, and Influence on Genetic Diversity
3.4. Approximate Bayesian Computation (ABC) Estimate of Divergence Time
4. Discussion
4.1. General Patterns of Genetic, Morphological, and Environmental Variation
4.2. Possible Implications for Conservation and Taxonomy
4.3. Caveats
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Source [Citation] | Fluorescent Dye | Repeat Unit |
|---|---|---|---|
| QS03797 | Chatwin et al. 2014 [98] | 6-FAM | (CA)7 |
| MSQ13 | Dow et al. 1995 [99] | 6-FAM | (TC)14 |
| QrZAG20 | Kampfer et al. 2004 [100] | VIC | (TC)18 |
| QpZAG110 | Steinkellner et al. 1997 [101] | NED | (AG)15 |
| QS00314 | Chatwin et al. 2014 [98] | PET | (GAA)6 |
| QS1904 | Chatwin et al. 2014 [98] | 6-FAM | (TC)10 |
| MSQ4 | Dow et al. 1995 [99] | VIC | (AG)17 |
| QS00562 | Steinkellner et al. 1997 [101] | 6-FAM | (GA)7 |
| QrZAG87 | Kampfer et al. 2004 [100] | NED | (TC)20 |
| QpZAG1.5 | Steinkellner et al. 1997 [101] | PET | (GT)5(GA)9 |
| QM69.2M1 | Isagi and Suhandono 1997 [102] | PET | (TGG)6(CGG)(TGG)2 |
| Parameter | East Mean | West Mean | T-test p-Value | Wilcoxon p-Value |
|---|---|---|---|---|
| Allelic richness | 47.160 | 58.877 | 0.0003 | 0.0013 |
| Heterozygosity | 0.587 | 0.653 | 0.0150 | 0.0148 |
| Pairwise FST | 0.085 | 0.079 | 0.3263 | 0.1111 |
| Relatedness | 0.176 | 0.077 | 0.0063 | 0.0093 |


| Eigenvalue | Variance (%) | Cumulative Variance (%) | |
|---|---|---|---|
| PC1 | 7.039 | 41.4 | 41.4 |
| PC2 | 3.584 | 21.1 | 62.5 |
| PC3 | 3.290 | 19.4 | 81.8 |
| PC4 | 1.459 | 8.6 | 90.4 |
| PC5 | 0.679 | 4.0 | 94.4 |
| PC6 | 0.494 | 2.9 | 97.3 |
| PC7 | 0.178 | 1.0 | 98.4 |
| PC8 | 0.093 | 0.5 | 98.9 |
| PC9 | 0.075 | 0.4 | 99.4 |
| PC10 | 0.045 | 0.3 | 99.6 |
| PC11 | 0.032 | 0.2 | 99.8 |
| PC12 | 0.021 | 0.1 | 99.9 |
| PC13 | 0.007 | 0.0 | 100.0 |
| PC14 | 0.003 | 0.0 | 100.0 |
| PC15 | 0.001 | 0.0 | 100.0 |
| PC16 | 0.000 | 0.0 | 100.0 |
| PC17 | 0.000 | 0.0 | 100.0 |
| Character | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Lobes | 0.32 | 0.07 | −0.20 | 0.19 |
| Length | −0.35 | 0.08 | −0.18 | −0.08 |
| BLBW | −0.26 | −0.21 | −0.25 | 0.09 |
| MLBW | 0.08 | −0.15 | −0.51 | −0.12 |
| ALBW | −0.22 | −0.14 | −0.02 | −0.59 |
| LVL | −0.31 | 0.18 | −0.19 | −0.04 |
| UVL | −0.04 | 0.19 | −0.49 | −0.05 |
| UMLS | −0.08 | −0.43 | −0.22 | −0.19 |
| LMLS | −0.20 | −0.37 | −0.20 | 0.12 |
| LLA | 0.35 | −0.07 | −0.10 | −0.15 |
| ULA | −0.19 | 0.31 | 0.16 | −0.17 |
| MLA | 0.26 | 0.09 | −0.06 | −0.41 |
| LWR_1 | 0.20 | −0.36 | 0.01 | 0.25 |
| LWR_2 | 0.35 | −0.17 | −0.11 | 0.03 |
| LWR_3 | 0.20 | −0.23 | 0.25 | −0.42 |
| LODR_1 | 0.17 | 0.33 | −0.28 | 0.11 |
| LODR_2 | 0.24 | 0.29 | −0.22 | −0.24 |
| Character | Abbreviation | CV1 | CV2 |
|---|---|---|---|
| Lobe Number | LOBES | −0.332 | −0.068 |
| Leaf Length | LENGTH | 0.052 | 0.025 |
| Basal Lobe Blade Width | BLBW | 0.018 | −0.300 |
| Middle Lobe Blade Width | MLBW | −0.135 | 0.299 |
| Apical Lobe Blade Width | ALBW | 0.058 | −0.392 |
| Lower Vein Length | LVL | 0.041 | −0.057 |
| Upper Vein Length | UVL | 0.018 | −0.084 |
| Upper Middle Lobe Sinus Width | UMLS | 0.008 | 0.174 |
| Lower Middle Lobe Sinus Width | LMLS | 0.065 | −0.075 |
| Lower Lobe Angle | LLA | −0.034 | 0.007 |
| Upper Lobe Angle | ULA | 0.014 | 0.013 |
| Middle Lobe Angle | MLA | 0.001 | −0.013 |
| Length to Width Ratio 1 (BLBW/Length) | LWR_1 | −0.332 | −0.068 |
| Length to Width Ratio 2 (MLBW/Length) | LWR_2 | 0.052 | 0.025 |
| Length to Width Ratio 3 (ALBW/Length) | LWR_3 | 0.018 | −0.300 |
| Lobe Depth Ratio 1 | LODR_1 | −0.135 | 0.299 |
| Lobe Depth Ratio 2 | LODR_2 | 0.058 | −0.392 |




| Bioclimatic Variable | Code | Range-Wide MLR and PCA | East MLR | West MLR | Niche Overlap |
|---|---|---|---|---|---|
| Annual Mean Temperature | BIO1 | ||||
| Mean Diurnal Range | BIO2 | X | X | ||
| Isothermality | BIO3 | X | |||
| Temperature Seasonality | BIO4 | X | |||
| Maximum Temperature of Warmest Month | BIO5 | X | X | ||
| Minimum Temperature of Coldest Month | BIO6 | ||||
| Temperature Annual Range | BIO7 | X | X | X | |
| Mean Temperature of Wettest Quarter | BIO8 | X | X | X | |
| Mean Temperature of Driest Quarter | BIO9 | X | X | X | |
| Mean Temperature of Warmest Quarter | BIO10 | X | |||
| Mean Temperature of Coldest Quarter | BIO11 | ||||
| Annual Precipitation | BIO12 | X | |||
| Precipitation of Wettest Month | BIO13 | ||||
| Precipitation of Driest Month | BIO14 | X | X | X | |
| Precipitation Seasonality | BIO15 | X | X | ||
| Precipitation of Wettest Quarter | BIO16 | ||||
| Precipitation of Driest Quarter | BIO17 | ||||
| Precipitation of Warmest Quarter | BIO18 | X | X | ||
| Precipitation of Coldest Quarter | BIO19 | X | X |
| Pop | BIO 1 | BIO2 | BIO3 | BIO4 | BIO5 | BIO6 | BIO7 | BIO8 | BIO9 | BIO10 | BIO 11 | BIO 12 | BIO 13 | BIO 14 | BIO 15 | BIO 16 | BIO 17 | BIO 18 | BIO 19 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E1 | 15 | 17.1 | 44 | 801.7 | 34.1 | −4.8 | 38.9 | 23.8 | 5 | 24.7 | 5 | 463 | 69 | 12 | 61.6 | 201 | 42 | 198 | 42 |
| E2 | 15.1 | 17.2 | 43.8 | 801.9 | 34.3 | −4.9 | 39.2 | 24.1 | 5.1 | 24.8 | 5.1 | 385 | 70 | 8 | 68.8 | 190 | 31 | 179 | 31 |
| E3 | 14.2 | 17.7 | 44.8 | 797.2 | 33.5 | −5.9 | 39.4 | 23.8 | 5.9 | 23.8 | 4.3 | 412 | 74 | 10 | 69.9 | 199 | 32 | 199 | 32 |
| E4 | 14.3 | 17.9 | 45.1 | 798.2 | 33.8 | −5.9 | 39.7 | 23.9 | 4.3 | 23.9 | 4.3 | 421 | 74 | 9 | 69 | 200 | 32 | 200 | 32 |
| E5 | 14.5 | 15 | 36.8 | 929.4 | 34.7 | −5.9 | 40.7 | 23.5 | 2.9 | 25.9 | 2.9 | 528 | 89 | 9 | 58.1 | 219 | 42 | 195 | 42 |
| E6 | 14.6 | 14.8 | 37 | 906 | 34.6 | −5.4 | 40 | 23.4 | 3.3 | 25.7 | 3.3 | 568 | 93 | 13 | 56 | 228 | 49 | 198 | 49 |
| E7 | 14.5 | 14.5 | 35.9 | 927.2 | 34.6 | −5.9 | 40.5 | 18.9 | 2.9 | 25.8 | 2.9 | 570 | 96 | 12 | 54.8 | 229 | 51 | 193 | 51 |
| E10 | 17.5 | 14.8 | 39.7 | 808.5 | 35.6 | −1.6 | 37.2 | 26.5 | 7.1 | 27 | 7.1 | 492 | 87 | 16 | 55.6 | 184 | 53 | 157 | 53 |
| E13 | 15.9 | 14.7 | 39.5 | 820.7 | 34.1 | −3 | 37.1 | 24.1 | 5.6 | 25.8 | 5.6 | 541 | 78 | 14 | 55.1 | 208 | 52 | 203 | 52 |
| E15 | 16.4 | 15.1 | 39.6 | 837 | 35.3 | −2.7 | 38.1 | 24.7 | 5.9 | 26.6 | 5.9 | 543 | 79 | 17 | 53 | 201 | 57 | 196 | 57 |
| W1 | 17.1 | 19.7 | 47.6 | 793.9 | 37.8 | −3.5 | 41.3 | 26.7 | 20.2 | 27.1 | 7.7 | 377 | 53 | 7 | 41.3 | 137 | 38 | 103 | 99 |
| W2 | 12.2 | 17.1 | 38.9 | 956 | 33.1 | −10.9 | 44 | 19.1 | 16.5 | 24.1 | 0.5 | 214 | 27 | 6 | 29.4 | 72 | 32 | 51 | 51 |
| W3 | 11.8 | 15.9 | 38.7 | 906.2 | 32.1 | −9.1 | 41.1 | 22.4 | 15.8 | 23.3 | 0.9 | 233 | 31 | 7 | 34.3 | 82 | 32 | 65 | 53 |
| W4 | 12.3 | 16.4 | 40.4 | 876.8 | 32.4 | −8.2 | 40.6 | 22.7 | 16.1 | 23.5 | 1.9 | 262 | 35 | 8 | 32.4 | 84 | 35 | 68 | 69 |
| W5 | 12.1 | 16 | 39.1 | 903.7 | 32.4 | −8.6 | 41 | 22.7 | 16.2 | 23.5 | 1.3 | 229 | 32 | 7 | 33.1 | 79 | 32 | 64 | 54 |
| W6 | 12.3 | 14.2 | 35.1 | 943.8 | 32.1 | −8.3 | 40.3 | 19 | 16.5 | 24.2 | 1 | 208 | 26 | 7 | 32.5 | 73 | 30 | 55 | 45 |
| W7 | 11.9 | 13.2 | 34 | 926.1 | 30.8 | −8 | 38.7 | 18.6 | 15.9 | 23.6 | 0.8 | 233 | 28 | 8 | 29.8 | 79 | 34 | 60 | 56 |
| W8 | 12.2 | 16 | 36.3 | 996.7 | 33.2 | −10.7 | 43.9 | 19 | −0.1 | 24.6 | −0.1 | 179 | 22 | 7 | 31.3 | 64 | 33 | 47 | 33 |
| W9 | 12.6 | 16.3 | 36.5 | 1002.6 | 33.7 | −10.8 | 44.5 | 19.6 | 22.3 | 25.1 | 0.2 | 245 | 31 | 9 | 24.9 | 74 | 51 | 53 | 52 |
| W10 | 12 | 17 | 36.3 | 1041.7 | 33.8 | −13.1 | 46.9 | 19.2 | −1.1 | 24.8 | −1.1 | 226 | 29 | 9 | 25.3 | 71 | 47 | 50 | 47 |
| W11 | 12.5 | 19.4 | 45.6 | 847.6 | 33.4 | −9.1 | 42.5 | 22.3 | 7.7 | 23.1 | 2.2 | 239 | 48 | 10 | 62.9 | 120 | 32 | 98 | 35 |
| W12 | 9.1 | 15 | 41.7 | 784.5 | 27.4 | −8.6 | 35.9 | 19.3 | 0 | 19.3 | 0 | 364 | 65 | 9 | 58.2 | 161 | 33 | 161 | 33 |
| WAUX3 | 12.3 | 16.2 | 36.5 | 1003.8 | 33.6 | −10.8 | 44.4 | 19.1 | 0 | 24.8 | 0 | 177 | 21 | 7 | 29.9 | 62 | 33 | 46 | 33 |
| East Mean | 15.2 | 15.9 | 40.6 | 842.8 | 34.5 | −4.6 | 39.1 | 23.7 | 4.8 | 25.4 | 4.6 | 492.3 | 80.9 | 12.0 | 60.2 | 205.9 | 44.1 | 191.8 | 44.1 |
| East ± SD | 1.1 | 1.4 | 3.5 | 55.5 | 0.7 | 1.6 | 1.3 | 1.9 | 1.4 | 1.1 | 1.5 | 68.2 | 9.7 | 3.1 | 6.7 | 15.1 | 9.7 | 13.9 | 9.7 |
| West Mean | 12.3 | 16.3 | 39.0 | 921.8 | 32.8 | −9.2 | 41.9 | 20.7 | 11.2 | 23.9 | 1.2 | 245.1 | 34.5 | 7.8 | 35.8 | 89.1 | 35.5 | 70.8 | 50.8 |
| West ± SD | 1.7 | 1.8 | 4 | 80.8 | 2.3 | 2.3 | 2.9 | 2.4 | 8.6 | 1.8 | 2.2 | 60.6 | 13 | 1.2 | 11.8 | 30.5 | 6.3 | 32.5 | 18.1 |
| Bioclimatic Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| BIO2: Mean Diurnal Range | −0.113 | −0.268 | 0.331 |
| BIO5: Max Temp of Warmest Month | 0.410 | 0.062 | 0.663 |
| BIO7: Annual Temp Range | 0.101 | −0.635 | 0.375 |
| BIO9: Mean Temp of Driest Quarter | 0.623 | −0.002 | −0.389 |
| BIO14: Precipitation of Driest Month | −0.334 | 0.610 | 0.364 |
| BIO19: Precipitation of Coldest Quarter | 0.556 | 0.385 | 0.164 |
| Eigenvalue | Variance (%) | Cumulative Variance (%) | |
|---|---|---|---|
| PC1 | 1.852 | 30.9 | 30.9 |
| PC2 | 1.542 | 25.7 | 56.6 |
| PC3 | 1.322 | 22.0 | 78.6 |
| PC4 | 0.921 | 15.3 | 94.0 |
| PC5 | 0.275 | 4.6 | 98.5 |
| PC6 | 0.088 | 1.5 | 100.0 |

| Model | Parameter | 0.025 CI | Median | 0.975 CI |
|---|---|---|---|---|
| IM | Theta 1 | 0.396 | 2.17 | 16.9 |
| IM | Theta 2 | 2.35 | 4.1 | 75.5 |
| IM | M | 1.75 × 10−10 | 0.177 | 5.52 |
| IM | TS | 0.0157 | 0.181 | 76.7 |
| I | Theta 1 | 0.1816 | 2.105 | 40.96 |
| I | Theta 2 | 1.938 | 3.47 | 10.43 |
| I | M | 1.75 × 10−10 | 0.1766 | 5.518 |
| I | TS | 0.003393 | 0.07413 | 0.3564 |
| Model | Parameter | 0.025 CI | Median | 0.975 CI |
|---|---|---|---|---|
| IM | Theta 1 | 157 | 863 | 6710 |
| IM | Theta 2 | 930 | 1630 | 29,900 |
| IM | M | 5.07 × 10−10 | 5.12 × 10−5 | 1.60 × 10−3 |
| IM | TS | 2710 | 31,100 | 13,200,000 |
| I | Theta 1 | 72 | 835 | 16,300 |
| I | Theta 2 | 769 | 1377 | 4140 |
| I | M | 5.23 × 10−10 | 5.29 × 10−5 | 1.65 × 10−3 |
| I | TS | 567 | 12,380 | 59,500 |
References
- Shafer, A.B.A.; Wolf, J.B.W. Widespread Evidence for Incipient Ecological Speciation: A Meta-Analysis of Isolation-by-Ecology. Ecol. Lett. 2013, 16, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A. Speciation in a Small Space. Proc. Natl. Acad. Sci. USA 2011, 108, 12975–12976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slatkin, M. Gene Flow and the Geographic Structure of Natural Populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Elam, D.R. Population Genetic Consequences of Small Population Size: Implications for Plant Conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]
- Jain, S.K.; Bradshaw, A.D. Evolutionary Divergence among Adjacent Plant Populations I. The Evidence and Its Theoretical Analysis. Heredity 1966, 21, 407–441. [Google Scholar] [CrossRef] [Green Version]
- Schluter, D. Ecology and the Origin of Species. Trends Ecol. Evol. 2001, 16, 372–380. [Google Scholar] [CrossRef]
- Rundle, H.D.; Nosil, P. Ecological Speciation. Ecol. Lett. 2005, 8, 336–352. [Google Scholar] [CrossRef]
- Thorne, R.F. Major Disjunctions in the Geographic Ranges of Seed Plants. Q. Rev. Biol. 1972, 47, 365–411. [Google Scholar] [CrossRef]
- Honnay, O.; Verheyen, K.; Butaye, J.; Jacquemyn, H.; Bossuyt, B.; Hermy, M. Possible Effects of Habitat Fragmentation and Climate Change on the Range of Forest Plant Species. Ecol. Lett. 2002, 5, 525–530. [Google Scholar] [CrossRef]
- Beatty, G.E.; Provan, J. Post-Glacial Dispersal, Rather than in Situ Glacial Survival, Best Explains the Disjunct Distribution of the Lusitanian Plant Species Daboecia cantabrica (Ericaceae). J. Biogeogr. 2012, 40, 335–344. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post-Glacial Re-Colonization of European Biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Zhao, M.; Chang, Y.; Kimball, R.T.; Zhao, J.; Lei, F.; Qu, Y. Pleistocene Glaciation Explains the Disjunct Distribution of the Chestnut-Vented Nuthatch (Aves, Sittidae). Zool. Scr. 2018, 48, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Grant, V. Plant Speciation, 2nd ed.; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Ortego, J.; Riordan, E.C.; Gugger, P.F.; Sork, V.L. Influence of Environmental Heterogeneity on Genetic Diversity and Structure in an Endemic Southern Californian Oak. Mol. Ecol. 2012, 21, 3210–3223. [Google Scholar] [CrossRef] [PubMed]
- Ülker, E.D.; Tavşanoğlu, Ç.; Perktaş, U. Ecological Niche Modelling of Pedunculate Oak (Quercus robur) Supports the ‘Expansion–Contraction’Model of Pleistocene Biogeography. Biol. J. Linn. Soc. 2018, 123, 338–347. [Google Scholar] [CrossRef]
- Di Pietro, R.; Conte, A.L.; Di Marzio, P.; Fortini, P.; Farris, E.; Gianguzzi, L.; Müller, M.; Rosati, L.; Spampinato, G.; Gailing, O. Does the Genetic Diversity among Pubescent White Oaks in Southern Italy, Sicily and Sardinia Islands Support the Current Taxonomic Classification? Eur. J. For. Res. 2020, 140, 355–371. [Google Scholar] [CrossRef]
- Fortini, P.; Antonecchia, G.; Di Marzio, P.; Maiuro, L.; Viscosi, V. Role of Micromorphological Leaf Traits and Molecular Data in Taxonomy of Three Sympatric White Oak Species and Their Hybrids (Quercus L.). Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2015, 149, 546–558. [Google Scholar]
- Fortini, P.; Di Marzio, P.; Di Pietro, R. Differentiation and Hybridization of Quercus frainetto, Q. petraea, and Q. pubescens, (Fagaceae): Insights from Macro-Morphological Leaf Traits and Molecular Data. Plant Syst. Evol. 2015, 301, 375–385. [Google Scholar] [CrossRef]
- Fortini, P.; Viscosi, V.; Maiuro, L.; Fineschi, S.; Vendramin, G. Comparative Leaf Surface Morphology and Molecular Data of Five Oaks of the Subgenus Quercus Oerst (Fagaceae). Plant Biosyst. 2009, 143, 543–554. [Google Scholar] [CrossRef]
- Manos, P.S.; Doyle, J.J.; Nixon, K.C. Phylogeny, Biogeography, and Processes of Molecular Differentiation in Quercus Subgenus Quercus (Fagaceae). Mol. Phylogenetics Evol. 1999, 12, 333–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavender-Bares, J.; González-Rodríguez, A.; Eaton, D.A.; Hipp, A.A.; Beulke, A.; Manos, P.S. Phylogeny and Biogeography of the American Live Oaks (Quercus Subsection Virentes): A Genomic and Population Genetics Approach. Mol. Ecol. 2015, 24, 3668–3687. [Google Scholar] [CrossRef]
- Hipp, A.L.; Manos, P.S.; González-Rodríguez, A.; Hahn, M.; Kaproth, M.; McVay, J.D.; Avalos, S.V.; Cavender-Bares, J. Sympatric Parallel Diversification of Major Oak Clades in the Americas and the Origins of Mexican Species Diversity. New Phytol. 2017, 217, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavender-Bares, J. Diversification, Adaptation, and Community Assembly of the American Oaks (Quercus), a Model Clade for Integrating Ecology and Evolution. New Phytol. 2018, 221, 669–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulos, H.M. A Review of the Evidence for Pine-Oak Niche Differentiation in the American Southwest. J. Sustain. For. 2009, 28, 92–107. [Google Scholar] [CrossRef]
- Felger, R.S.; Johnson, M.B. Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and Northwestern Mexico. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and northern Mexico; The Station: Fort Collins, CO, USA, 1995; Volume 264, pp. 71–77. [Google Scholar]
- Dhillion, S.S.; Mills, M.H. The sand shinnery oak (Quercus havardii) communities of the Llano Estacado: History, structure, ecology, and restoration. In Savannas, Barrens, and Rock Outcrop Plant Communities of North America; Cambridge University Press: Cambridge, UK, 1999; pp. 262–274. [Google Scholar]
- Peterson, R.; Boyd, C.S. Ecology and Management of Sand Shinnery Communities: A Literature Review; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000.
- Dhillion, S.S. Environmental Heterogeneity, Animal Disturbances, Microsite Characteristics, and Seedling Establishment in a Quercus havardii Community. Restor. Ecol. 1999, 7, 399–406. [Google Scholar] [CrossRef]
- Davis, W.J. Shin-Oak (Quercus havardii, Rydb.; Fagaceae) Rhizome Shoot Production: Possibilities for Use in Restoration. M.S.; Texas Tech University: Lubbock, TX, USA, 2013. [Google Scholar]
- Jerome, D.; Beckman, E.; Kenny, L.; Wenzell, K.; Kua, C.; Westwood, M. The Red List of US Oaks; The Morton Arboretum: Lisle, IL, USA, 2017. [Google Scholar]
- Tucker, J.M. Studies in the Quercus undulata Complex. IV. The Contribution of Quercus havardii. Am. J. Bot. 1970, 57, 71–84. [Google Scholar] [CrossRef]
- McCauley, R.A.; Christie, B.J.; Ireland, E.L.; Landers, R.A.; Nichols, H.R.; Schendel, M.T. Influence of Relictual Species on the Morphology of a Hybridizing Oak Complex: An Analysis of the Quercus x undulata Complex in the Four Corners Region. West. N. Am. Nat. 2012, 72, 296–310. [Google Scholar] [CrossRef]
- Jarčuška, B.; Milla, R. Shoot-Level Biomass Allocation Is Affected by Shoot Type in Fagus sylvatica. J. Plant Ecol. 2012, 5, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Cicák, A. Estimation of Morphological Parameters for European Beech Leaves on Spring Shoots Using the Method of Calculation Coefficients. Oecol 2003, 30, 131–140. [Google Scholar]
- Viscosi, V.; Lepais, O.; Gerber, S.; Fortini, P. Leaf Morphological Analyses in Four European Oak Species (Quercus) and Their Hybrids: A Comparison of Traditional and Geometric Morphometric Methods. Plant Biosyst. 2009, 143, 564–574. [Google Scholar] [CrossRef]
- Albarrán-Lara, A.L.; Mendoza-Cuenca, L.; Valencia-Avalos, S.; González-Rodríguez, A.; Oyama, K. Leaf Fluctuating Asymmetry Increases with Hybridization and Introgression between Quercus magnoliifolia and Quercus resinosa (Fagaceae) through an Altitudinal Gradient in Mexico. Int. J. Plant Sci. 2010, 171, 310–322. [Google Scholar] [CrossRef]
- Holmgren, P.K.; Holmgren, N.H. Index Herbariorum. Taxon 1991, 40, 687–692. [Google Scholar] [CrossRef]
- Holleley, C.E.; Geerts, P.G. Multiplex Manager 1.0: A Cross-Platform Computer Program That Plans and Optimizes Multiplex PCR. BioTechniques 2009, 46, 511–517. [Google Scholar] [CrossRef]
- Dzialuk, A.; Chybicki, I.; Burczyk, J. PCR Multiplexing of Nuclear Microsatellite Loci in Quercus Species. Plant Mol. Biol. Rep. 2005, 23, 121–128. [Google Scholar] [CrossRef]
- Abraham, S.T.; Zaya, D.N.; Koenig, W.D.; Ashley, M.V. Interspecific and Intraspecific Pollination Patterns of Valley Oak, Quercus lobata, in a Mixed Stand in Coastal Central California. Int. J. Plant Sci. 2011, 172, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Backs, J.R.; Terry, M.; Ashley, M.V. Using Genetic Analysis to Evaluate Hybridization as a Conservation Concern for the Threatened Species Quercus hinckleyi C.H. Muller (Fagaceae). Int. J. Plant Sci. 2016, 177, 121–131. [Google Scholar] [CrossRef]
- Craft, K.J.; Ashley, M.V. Landscape Genetic Structure of Bur Oak (Quercus macrocarpa) Savannas in Illinois. For. Ecol. Manag. 2007, 239, 13–20. [Google Scholar] [CrossRef]
- Oosterhout, C.V.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. Micro-Checker: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Chybicki, I.J.; Burczyk, J. Simultaneous Estimation of Null Alleles and Inbreeding Coefficients. J. Hered. 2008, 100, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R Package for Genetic Analysis of Populations with Clonal, Partially Clonal, and/or Sexual Reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T. Adegenet: A R Package for the Multivariate Analysis of Genetic Markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. Hierfstat, a Package for R to Compute and Test Hierarchical F-Statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, P.; Gerlach, G. Demerelate: Calculating Interindividual Relatedness for Kinship Analysis Based on Codominant Diploid Genetic Markers Using R. Mol. Ecol. Resour. 2017, 17, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research–an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Rosenberg, N.A. Distruct: A Program for the Graphical Display of Population Structure. Mol. Ecol. Notes 2003, 4, 137–138. [Google Scholar] [CrossRef]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant Analysis of Principal Components: A New Method for the Analysis of Genetically Structured Populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Uslu, E.; Bakış, Y. Morphometric Analyses of the Leaf Variation within Quercus L. Sect. Cerris Loudon in Turkey. Dendrobiology 2013, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.S. Variation of Growth in Danish Provenance Trials with Oak (Quercus robur L. and Quercus petraea Mattuschka Liebl.). Ann. Des Sci. For. 1993, 50, 203s–207s. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2012, 36, 27–46. [Google Scholar] [CrossRef]
- Chatterjee, S.; Hadi, A.S.; Price, B. Simple Linear Regression. In Regression Analysis by Example; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 21–51. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Linear and Nonlinear Mixed Effects Models. R package version 2007, 3, 1–89. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2012, 4, 133–142. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1. 0. 0. 2009. Available online: http://r-forge.r-project.org/projects/mumin/ (accessed on 13 March 2021).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Schoener, T.W. Nonsynchronous Spatial Overlap of Lizards in Patchy Habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Lumley, T. Based on R Code by A. M. Leaps: Regression Subset Selection, R Package Version 3.0. Available online: https://cran.r-project.org/package=leaps (accessed on 10 March 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Yang, J.-Q.; Hsu, K.-C.; Liu, Z.-Z.; Su, L.-W.; Kuo, P.-H.; Tang, W.-Q.; Zhou, Z.-C.; Liu, D.; Bao, B.-L.; Lin, H.-D. The Population History of Garra orientalis (Teleostei: Cyprinidae) Using Mitochondrial DNA and Microsatellite Data with Approximate Bayesian Computation. BMC Evol. Biol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-W.; Li, Y.; Zhang, Q.; Fang, Y.-M. Ancient East-West Divergence, Recent Admixture, and Multiple Marginal Refugia Shape Genetic Structure of a Widespread Oak Species (Quercus acutissima) in China. Tree Genet. Genomes 2018, 14, 88. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, Y.-P.; Chen, X.-D.; Yang, J.; Zhou, T.; Bai, G.-Q.; Yang, J.; Li, Z.-H.; Peng, C.-I.; Zhao, G.-F. Allopatric Divergence, Local Adaptation, and Multiple Quaternary Refugia in a Long-Lived Tree (Quercus spinosa) from Subtropical China. BioRxiv 2017. [Google Scholar] [CrossRef] [Green Version]
- Navascués, M. MicrosatABC-IM: Approximate Bayesian Computation Inferences under the Isolation with Migration Model from Microsatellite Data. Available online: https://hal.inrae.fr/hal-02789612 (accessed on 20 February 2021). [CrossRef]
- Hudson, R.R. Generating Samples under a Wright-Fisher Neutral Model of Genetic Variation. Bioinformatics 2002, 18, 337–338. [Google Scholar] [CrossRef]
- Raynal, L.; Marin, J.-M.; Pudlo, P.; Ribatet, M.; Robert, C.P.; Estoup, A. ABC Random Forests for Bayesian Parameter Inference. Bioinformatics 2018, 35, 1720–1728. [Google Scholar] [CrossRef]
- Pudlo, P.; Marin, J.-M.; Estoup, A.; Cornuet, J.-M.; Gautier, M.; Robert, C.P. Reliable ABC Model Choice via Random Forests. Bioinformatics 2016, 32, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Thuillet, A.-C.; Bru, D.; David, J.; Roumet, P.; Santoni, S.; Sourdille, P.; Bataillon, T. Direct Estimation of Mutation Rate for 10 Microsatellite Loci in Durum Wheat, Triticum turgidum (L.) Thell. ssp. Durum Desf. Mol. Biol. Evol. 2002, 19, 122–125. [Google Scholar] [CrossRef] [Green Version]
- Vigouroux, Y.; Jaqueth, J.S.; Matsuoka, Y.; Smith, O.S.; Beavis, W.D.; Smith, J.S.C.; Doebley, J. Rate and Pattern of Mutation at Microsatellite Loci in Maize. Mol. Biol. Evol. 2002, 19, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Marriage, T.N.; Hudman, S.; Mort, M.E.; Orive, M.E.; Shaw, R.G.; Kelly, J.K. Direct Estimation of the Mutation Rate at Dinucleotide Microsatellite Loci in Arabidopsis thaliana (Brassicaceae). Heredity 2009, 103, 310–317. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, L.M. Somatic Mutations at Microsatellite Loci in Western Red Cedar (Thuja plicata: Cupressaceae). J. Hered. 2004, 95, 172–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kass, R.E.; Raftery, A.E. Bayes Factors. J. Am. Stat. Assoc. 1995, 90, 773–795. [Google Scholar] [CrossRef]
- Jarosz, A.F.; Wiley, J. What Are the Odds? A Practical Guide to Computing and Reporting Bayes Factors. J. Probl. Solving 2014, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Ashley, M.V.; Backs, J.R.; Kindsvater, L.; Abraham, S.T. Genetic Variation and Structure in an Endemic Island Oak, Quercus tomentella, and Mainland Canyon Oak, Quercus chrysolepis. Int. J. Plant Sci. 2018, 179, 151–161. [Google Scholar] [CrossRef]
- Backs, J.R.; Terry, M.; Klein, M.; Ashley, M.V. Genetic Analysis of a Rare Isolated Species: A Tough Little West Texas Oak, Quercus hinckleyi C.H. Mull. J. Torrey Bot. Soc. 2015, 142, 302–313. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, A.; Oyama, K. Leaf Morphometric Variation in Quercus affinis and Q. laurina (Fagaceae), Two Hybridizing Mexican Red Oaks. Bot. J. Linn. Soc. 2005, 147, 427–435. [Google Scholar] [CrossRef]
- Kusi, J.; Karsai, I. Plastic Leaf Morphology in Three Species of Quercus: The More Exposed Leaves Are Smaller, More Lobated and Denser. Plant Species Biol. 2019, 35, 24–37. [Google Scholar] [CrossRef]
- Trejo, L.; Alvarado-Cárdenas, L.O.; Scheinvar, E.; Eguiarte, L.E. Population Genetic Analysis and Bioclimatic Modeling in Agave striata in the Chihuahuan Desert Indicate Higher Genetic Variation and Lower Differentiation in Drier and More Variable Environments. Am. J. Bot. 2016, 103, 1020–1029. [Google Scholar] [CrossRef] [Green Version]
- Sork, V.L.; Davis, F.W.; Westfall, R.; Flint, A.; Ikegami, M.; Wang, H.; Grivet, D. Gene Movement and Genetic Association with Regional Climate Gradients in California Valley Oak (Quercus lobata Née) in the Face of Climate Change. Mol. Ecol. 2010, 19, 3806–3823. [Google Scholar] [CrossRef]
- Riordan, E.C.; Gugger, P.F.; Ortego, J.; Smith, C.; Gaddis, K.; Thompson, P.; Sork, V.L. Association of Genetic and Phenotypic Variability with Geography and Climate in Three Southern California Oaks. Am. J. Bot. 2016, 103, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Vázquez, L.; Feng, L.; Liu, Z.; Zhao, G. Climatic and Soil Factors Shape the Demographical History and Genetic Diversity of a Deciduous Oak (Quercus liaotungensis) in Northern China. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef]
- Ortego, J.; Gugger, P.F.; Sork, V.L. Climatically Stable Landscapes Predict Patterns of Genetic Structure and Admixture in the Californian Canyon Live Oak. J. Biogeogr. 2014, 42, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Mayes, S.G.; McGinley, M.A.; Werth, C.R. Clonal Population Structure and Genetic Variation in Sand-Shinnery Oak, Quercus havardii (Fagaceae). Am. J. Bot. 1998, 85, 1609–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipp, A.L.; Eaton, D.A.R.; Cavender-Bares, J.; Fitzek, E.; Nipper, R.; Manos, P.S. A Framework Phylogeny of the American Oak Clade Based on Sequenced RAD Data. PLoS ONE 2014, 9, e93975. [Google Scholar] [CrossRef] [Green Version]
- Hampe, A.; Petit, R.J. Conserving Biodiversity under Climate Change: The Rear Edge Matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Muller, C.H. Ecological Control of Hybridization in Quercus: A Factor in the Mechanism of Evolution. Evolution 1952, 6, 147. [Google Scholar] [CrossRef]
- Hipp, A.L.; Manos, P.S.; Hahn, M.; Avishai, M.; Bodénès, C.; Cavender-Bares, J.; Crowl, A.A.; Deng, M.; Denk, T.; Fitz-Gibbon, S.; et al. Genomic Landscape of the Global Oak Phylogeny. New Phytol. 2019, 226, 1198–1212. [Google Scholar] [CrossRef]
- Lepais, O.; Gerber, S. Reproductive Patterns Shape Introgression Dynamics and Species Succession within the European White Oak Species Complex. Evolution 2010, 65, 156–170. [Google Scholar] [CrossRef]
- Spence, E.S.; Fant, J.; Gailing, O.; Griffith, M.P.; Havens, K.; Hipp, A.L.; Kadav, P.; Kramer, A.; Thompson, P.; Toppila, R.; et al. Comparing genetic diversity in three threatened oaks. Forests 2021, 12. [Google Scholar]
- Chatwin, W.B.; Carpenter, K.K.; Jimenez, F.R.; Elzinga, D.B.; Johnson, L.A.; Maughan, P.J. Microsatellite Primer Development for Post Oak, Quercus stellata (Fagaceae). Appl. Plant Sci. 2014, 2, 1400070. [Google Scholar] [CrossRef] [PubMed]
- Dow, B.D.; Ashley, M.V.; Howe, H.F. Characterization of Highly Variable (GA/CT) n Microsatellites in the Bur Oak, Quercus macrocarpa. Theor. Appl. Genet. 1995, 91, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, S.; Lexer, C.; Glössl, J.; Steinkellner, H. Characterization of (GA) n Microsatellite Loci from Quercus robur. Hereditas 2004, 129, 183–186. [Google Scholar] [CrossRef]
- Steinkellner, H.; Lexer, C.; Turetschek, E.; Glössl, J. Conservation of (GA) n Microsatellite Loci between Quercus Species. Mol. Ecol. 1997, 6, 1189–1194. [Google Scholar] [CrossRef]
- Isagi, Y.; Suhandono, S. PCR Primers Amplifying Microsatellite Loci of Quercus myrsinifolia Blume and Their Conservation between Oak Species. Mol. Ecol. 1997, 6, 897–899. [Google Scholar] [CrossRef] [PubMed]



| Population | Latitude (N) | Longitude (W) | n | MLG | Clones | He | Ho | Na | Ar | FST | Rel |
|---|---|---|---|---|---|---|---|---|---|---|---|
| E1 | 33.373 | −102.617 | 20 | 18 | 2 | 0.554 | 0.524 | 58 | 3.61 | 0.088 | 0.215 |
| E2 | 33.408 | −103.867 | 25 | 22 | 3 | 0.562 | 0.496 | 56 | 3.42 | 0.090 | 0.250 |
| E3 | 33.712 | −103.344 | 27 | 20 | 7 | 0.520 | 0.418 | 57 | 3.44 | 0.094 | 0.239 |
| E4 | 33.609 | −103.183 | 19 | 13 | 6 | 0.542 | 0.433 | 49 | 3.58 | 0.082 | 0.149 |
| E5 | 35.903 | −100.276 | 32 | 17 | 15 | 0.543 | 0.516 | 60 | 3.55 | 0.095 | 0.265 |
| E6 | 35.445 | −100.199 | 14 | 11 | 3 | 0.654 | 0.570 | 60 | 4.26 | 0.078 | 0.113 |
| E7 | 35.767 | −99.926 | 56 | 40 | 16 | 0.615 | 0.554 | 91 | 4.16 | 0.072 | 0.156 |
| E10 | 32.286 | −101.332 | 9 | 9 | 0 | 0.649 | 0.581 | 55 | 4.38 | 0.066 | 0.054 |
| E13 | 33.459 | −101.391 | 8 | 8 | 0 | 0.634 | 0.648 | 53 | 4.29 | 0.073 | 0.158 |
| E15 | 33.435 | −101.074 | 24 | 21 | 3 | 0.704 | 0.591 | 84 | 4.82 | 0.061 | 0.009 |
| W1 | 34.727 | −111.985 | 39 | 25 | 14 | 0.553 | 0.478 | 58 | 3.37 | 0.129 | 0.275 |
| W2 | 36.927 | −109.618 | 11 | 7 | 4 | 0.681 | 0.494 | 53 | 4.57 | 0.077 | −0.044 |
| W3 | 36.616 | −110.133 | 33 | 32 | 1 | 0.586 | 0.491 | 88 | 4.53 | 0.083 | 0.098 |
| W4 | 37.096 | −111.984 | 33 | 26 | 7 | 0.646 | 0.614 | 79 | 4.42 | 0.076 | 0.143 |
| W5 | 36.786 | −111.540 | 30 | 27 | 3 | 0.668 | 0.616 | 103 | 5.05 | 0.061 | 0.052 |
| W6 | 37.064 | −110.069 | 33 | 31 | 2 | 0.663 | 0.560 | 106 | 4.90 | 0.061 | 0.051 |
| W7 | 37.234 | −109.973 | 32 | 30 | 2 | 0.660 | 0.567 | 93 | 4.77 | 0.064 | 0.078 |
| W8 | 38.065 | −110.602 | 39 | 35 | 4 | 0.716 | 0.616 | 98 | 4.94 | 0.060 | 0.035 |
| W9 | 38.573 | −109.527 | 30 | 28 | 2 | 0.701 | 0.552 | 106 | 5.23 | 0.058 | −0.001 |
| W10 | 38.762 | −109.725 | 29 | 22 | 7 | 0.696 | 0.603 | 90 | 5.14 | 0.066 | 0.021 |
| W11 | 34.998 | −107.172 | 20 | 19 | 1 | 0.647 | 0.584 | 84 | 4.80 | 0.086 | 0.089 |
| W12 | 38.502 | −105.106 | 18 | 15 | 3 | 0.601 | 0.453 | 69 | 4.38 | 0.093 | 0.081 |
| WAUX3 | 38.000 | −110.569 | 16 | 14 | 2 | 0.694 | 0.641 | 73 | 4.86 | 0.070 | 0.051 |
| East Mean | 23.4 | 17.9 | 5.5 | 0.598 | 0.533 | 62.3 | 3.95 | 0.080 | 0.161 | ||
| West Mean | 27.9 | 23.9 | 4.0 | 0.655 | 0.559 | 84.6 | 4.69 | 0.076 | 0.072 | ||
| Overall Mean | 26.0 | 21.3 | 4.7 | 0.630 | 0.548 | 74.1 | 4.79 | 0.077 | 0.110 |
| Character | Abbreviation | Regional | Local |
|---|---|---|---|
| Lobe Number | LOBES | 0.182 | 0.345 |
| Leaf Length | LENGTH | 0.309 | 0.332 |
| Basal Lobe Blade Width | BLBW | 0.064 | 0.449 |
| Middle Lobe Blade Width | MLBW | 0.010 | 0.491 |
| Apical Lobe Blade Width | ALBW | 0.023 | 0.025 |
| Lower Vein Length | LVL | 0.105 | 0.145 |
| Upper Vein Length | UVL | 0.002 | 0.342 |
| Upper Middle Lobe Sinus Width | UMLS | 0.001 | 0.508 |
| Lower Middle Lobe Sinus Width | LMLS | 0.027 | 0.470 |
| Lower Lobe Angle | LLA | 0.383 | 0.202 |
| Upper Lobe Angle | ULA | 0.038 | 0.161 |
| Middle Lobe Angle | MLA | 0.038 | 0.205 |
| Length to Width Ratio 1 (BLBW/Length) | LWR_1 | 0.058 | 0.442 |
| Length to Width Ratio 2 (MLBW/Length) | LWR_2 | 0.403 | 0.264 |
| Length to Width Ratio 3 (ALBW/Length) | LWR_3 | 0.040 | 0.223 |
| Lobe Depth Ratio 1 | LODR_1 | 0.026 | 0.402 |
| Lobe Depth Ratio 2 | LODR_2 | 0.076 | 0.411 |
| Range-Wide MLR | East MLR | West MLR | |||||||
|---|---|---|---|---|---|---|---|---|---|
| He | Ar | Rel | He | Ar | Rel | He | Ar | Rel | |
| BIO2: Mean Diurnal Range | 0.0473 | 0.0013 | 0.0105 | 0.0049 | 0.0016 | 0.0194 | |||
| BIO3: Isothermality | 0.0118 | ||||||||
| BIO5: Max Temp of Warmest Month | 0.1034 | ||||||||
| BIO7: Annual Temp Range | 0.0002 | 0.0019 | 0.0153 | 0.0008 | |||||
| BIO8: Mean Temp of Wettest Quarter | 0.0004 | 0.0501 | 0.0011 | ||||||
| BIO9: Mean Temp of Driest Quarter | 0.0181 | 0.1306 | 0.0451 | 0.0775 | |||||
| BIO10: Mean Temp of Warmest Quarter | 0.0023 | ||||||||
| BIO12: Annual Precipitation | 0.0001 | 0.0635 | |||||||
| BIO14: Precipitation of Driest Month | 0.0041 | 0.1390 | 0.0006 | ||||||
| BIO15: Precipitation Seasonality | 0.0231 | 0.0002 | |||||||
| BIO18: Precipitation of Warmest Quarter | 0.1484 | ||||||||
| BIO19: Precipitation of Coldest Quarter | 0.0000 | ||||||||
| Model Adjusted R2 | 0.6121 | 0.5019 | 0.3457 | 0.6286 | 0.7527 | 0.5093 | 0.9303 | 0.9665 | 0.7257 |
| Model p-value | 0.0026 | 0.0007 | 0.0111 | 0.0259 | 0.0275 | 0.0687 | 0.0005 | 0.0003 | 0.0012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zumwalde, B.A.; McCauley, R.A.; Fullinwider, I.J.; Duckett, D.; Spence, E.; Hoban, S. Genetic, Morphological, and Environmental Differentiation of an Arid-Adapted Oak with a Disjunct Distribution. Forests 2021, 12, 465. https://doi.org/10.3390/f12040465
Zumwalde BA, McCauley RA, Fullinwider IJ, Duckett D, Spence E, Hoban S. Genetic, Morphological, and Environmental Differentiation of an Arid-Adapted Oak with a Disjunct Distribution. Forests. 2021; 12(4):465. https://doi.org/10.3390/f12040465
Chicago/Turabian StyleZumwalde, Bethany A., Ross A. McCauley, Ian J. Fullinwider, Drew Duckett, Emma Spence, and Sean Hoban. 2021. "Genetic, Morphological, and Environmental Differentiation of an Arid-Adapted Oak with a Disjunct Distribution" Forests 12, no. 4: 465. https://doi.org/10.3390/f12040465