

Boesenbergia stenophylla-Derived Stenophyllol B Exerts Antiproliferative and Oxidative Stress Responses in Triple-Negative Breast Cancer Cells with Few Side Effects in Normal Cells

 , , , and
, , , and 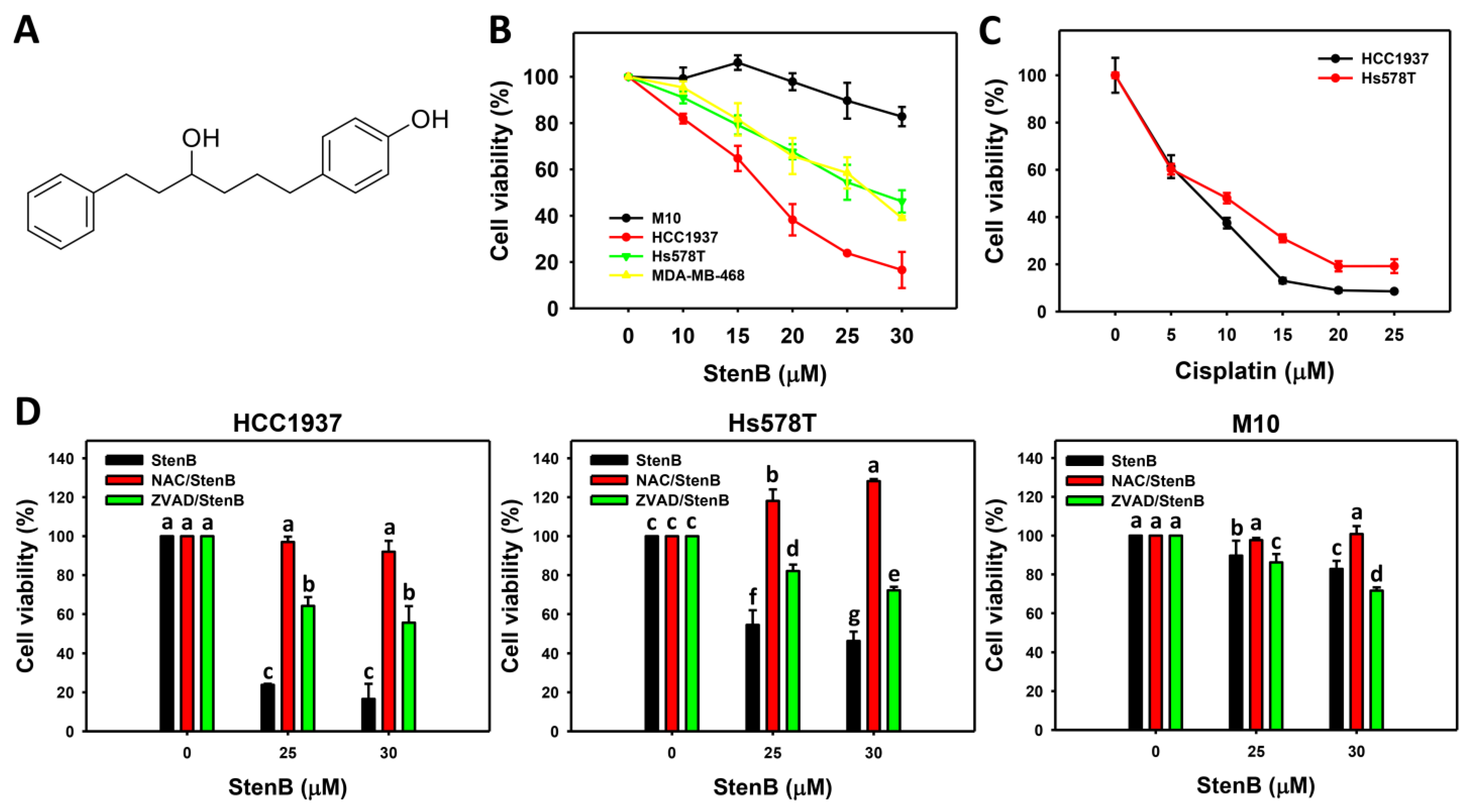{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Antiproliferation Response to StenB (TNBC vs. Normal Cells)
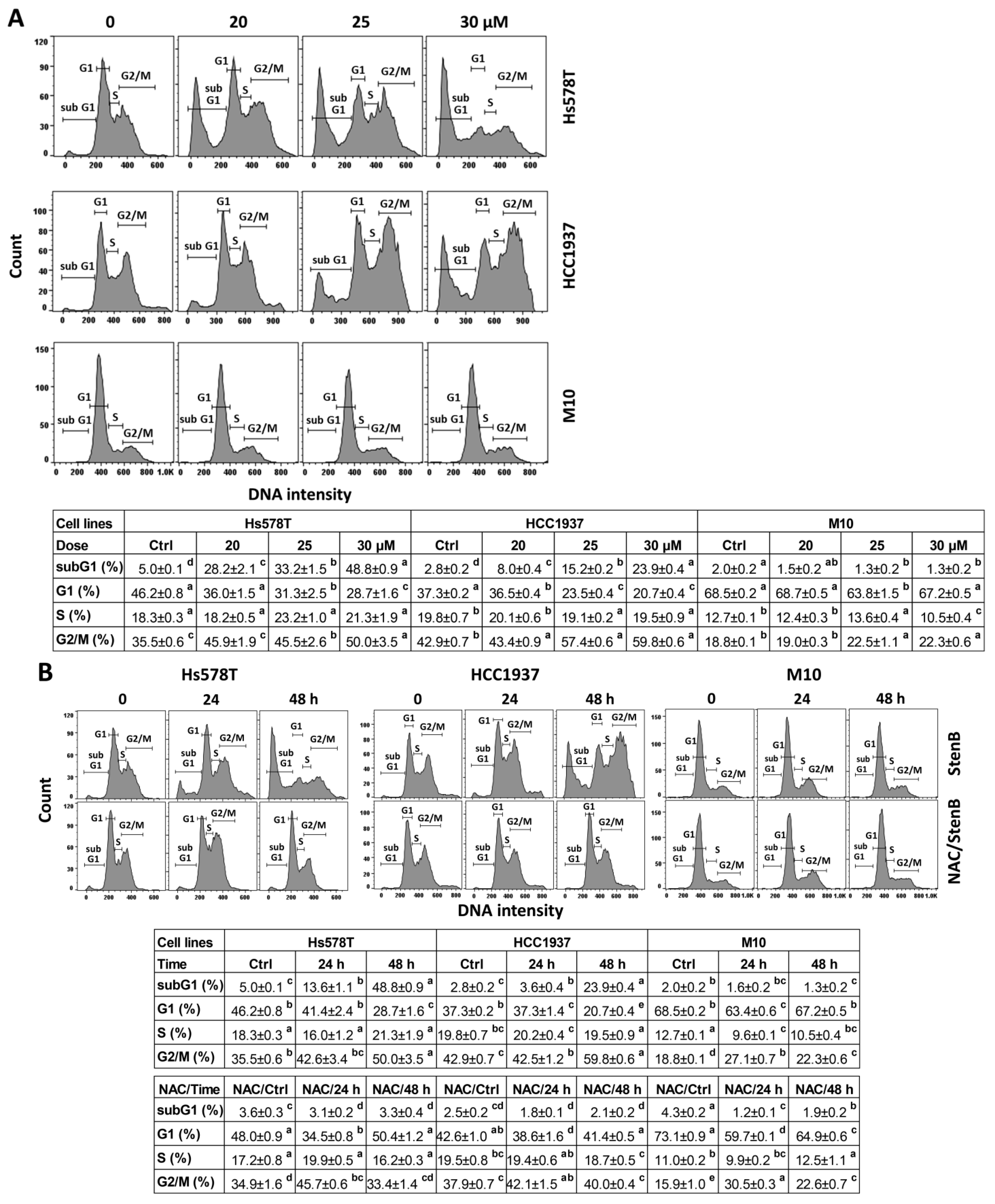
2.2. Cell Cycle Response to StenB (TNBC vs. Normal Cells)
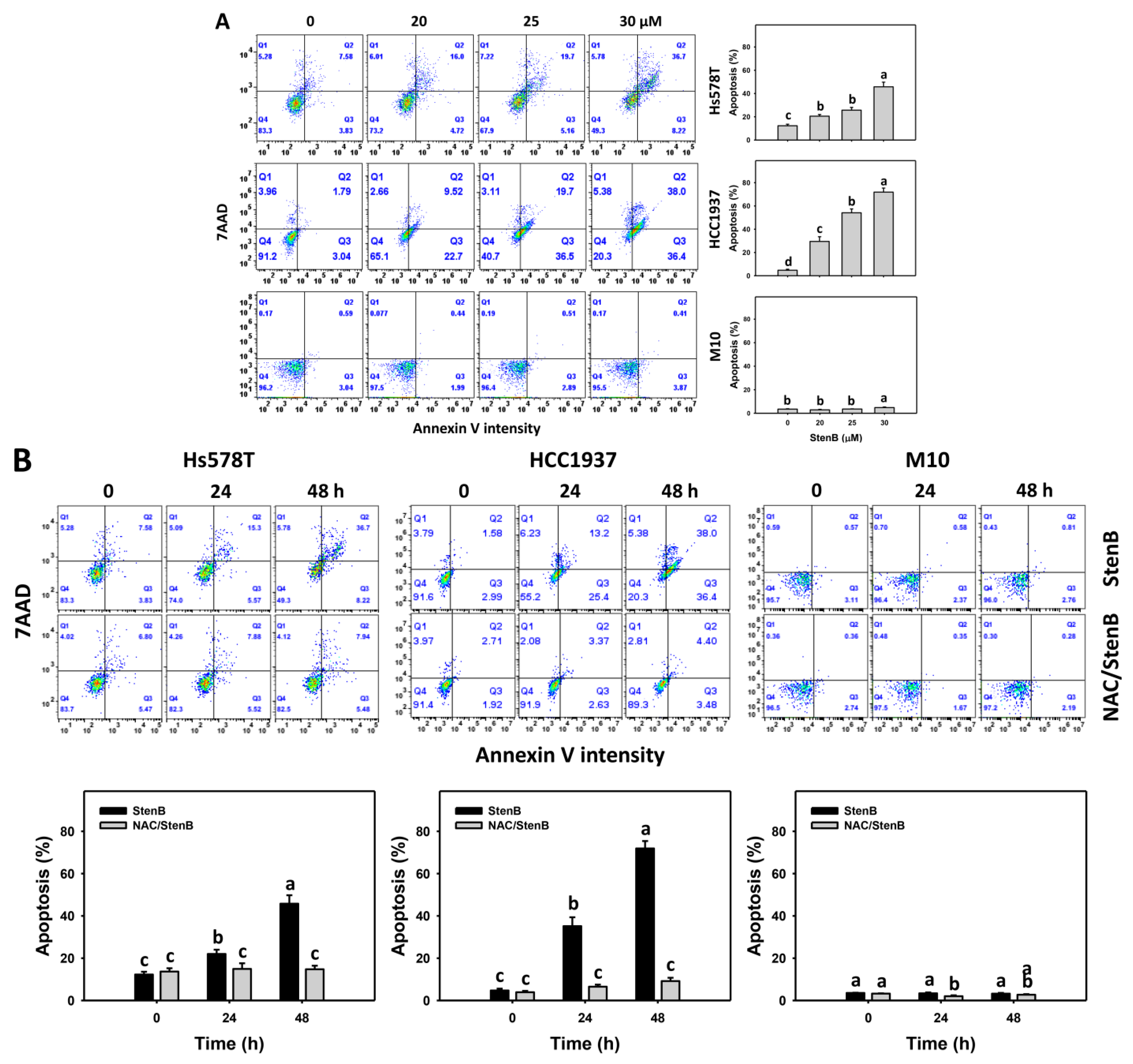
2.3. Annexin V Response to StenB (TNBC vs. Normal Cells)
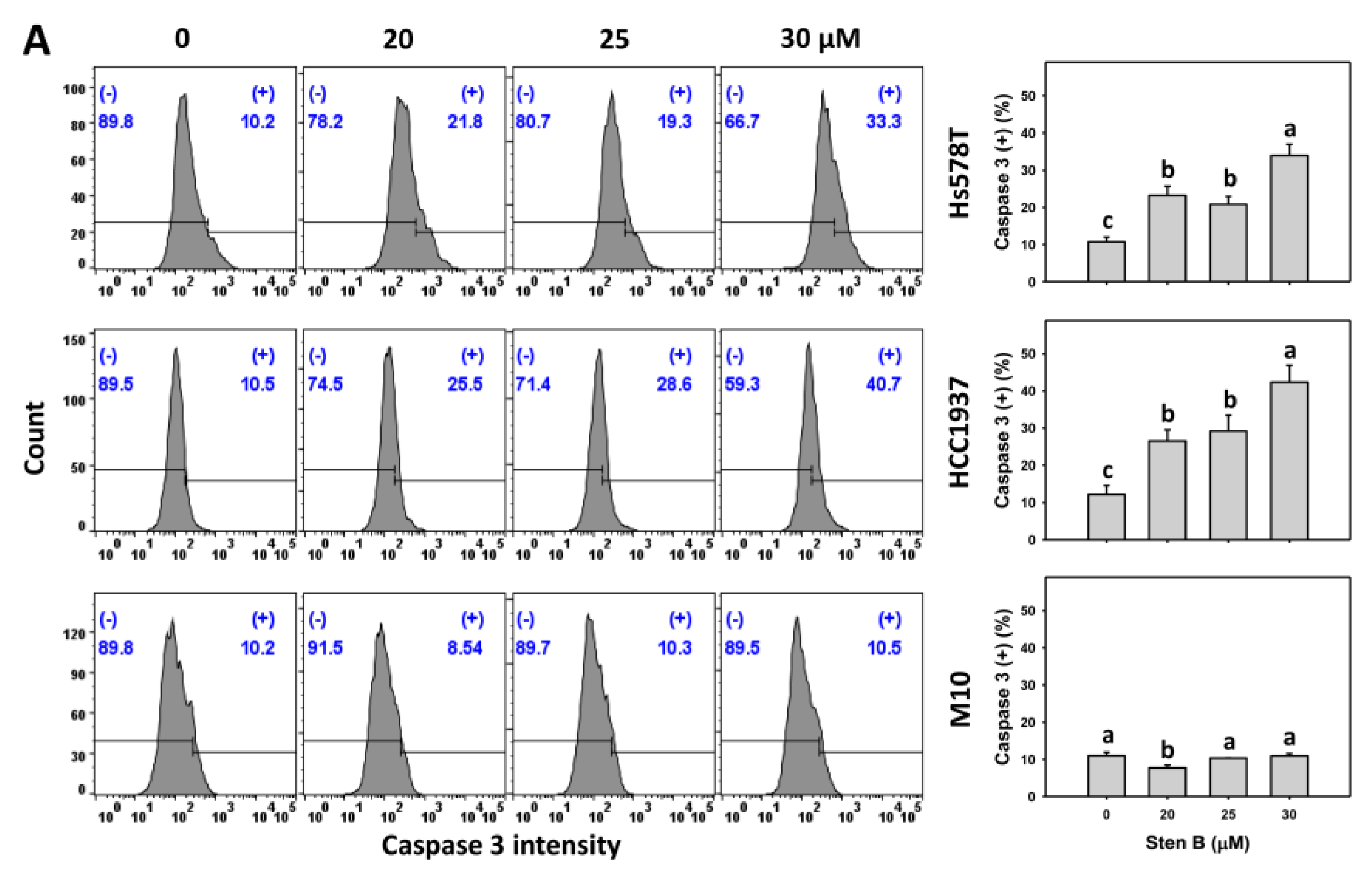
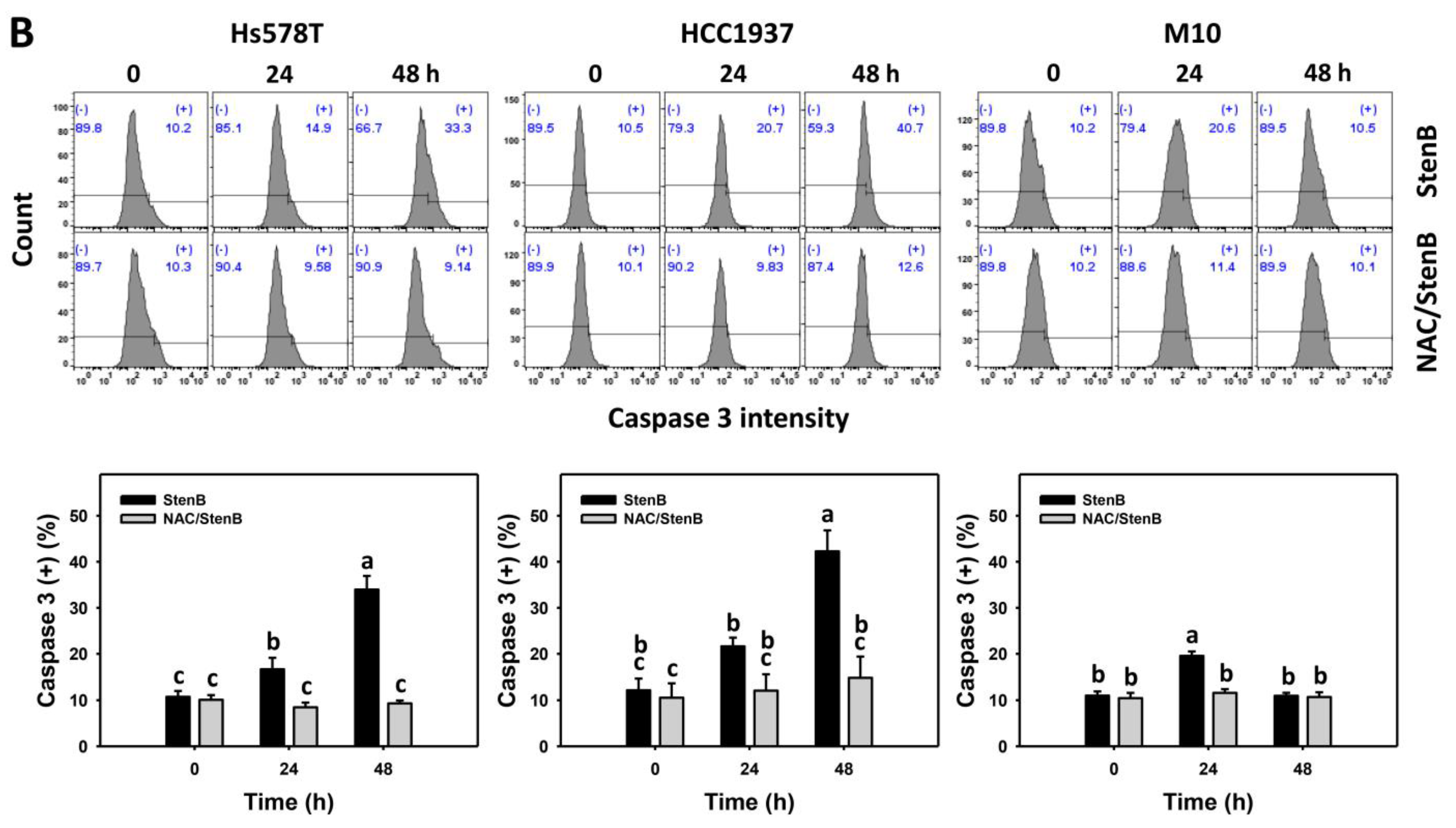
2.4. Caspase 3 Responses to StenB (TNBC vs. Normal Cells)
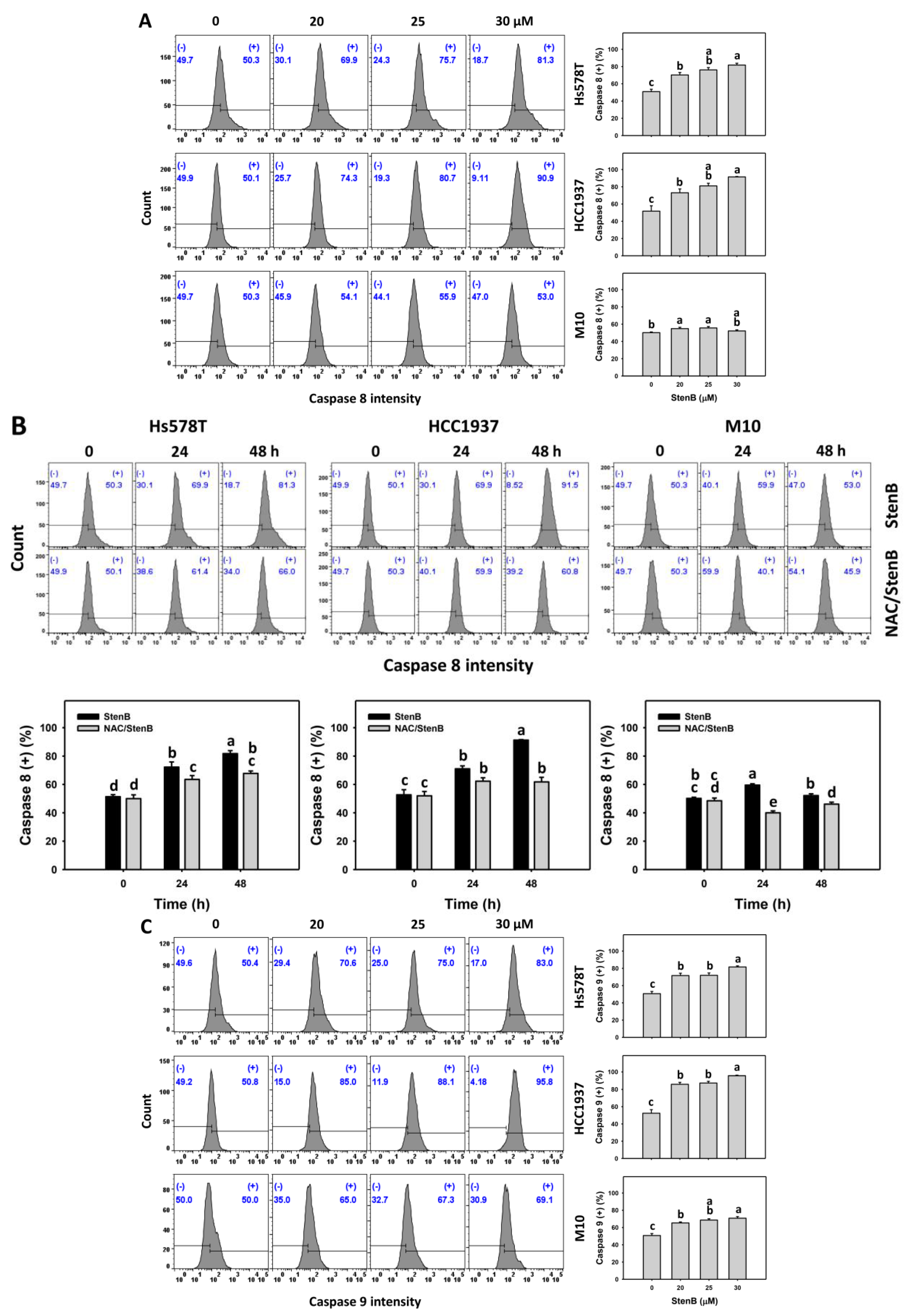
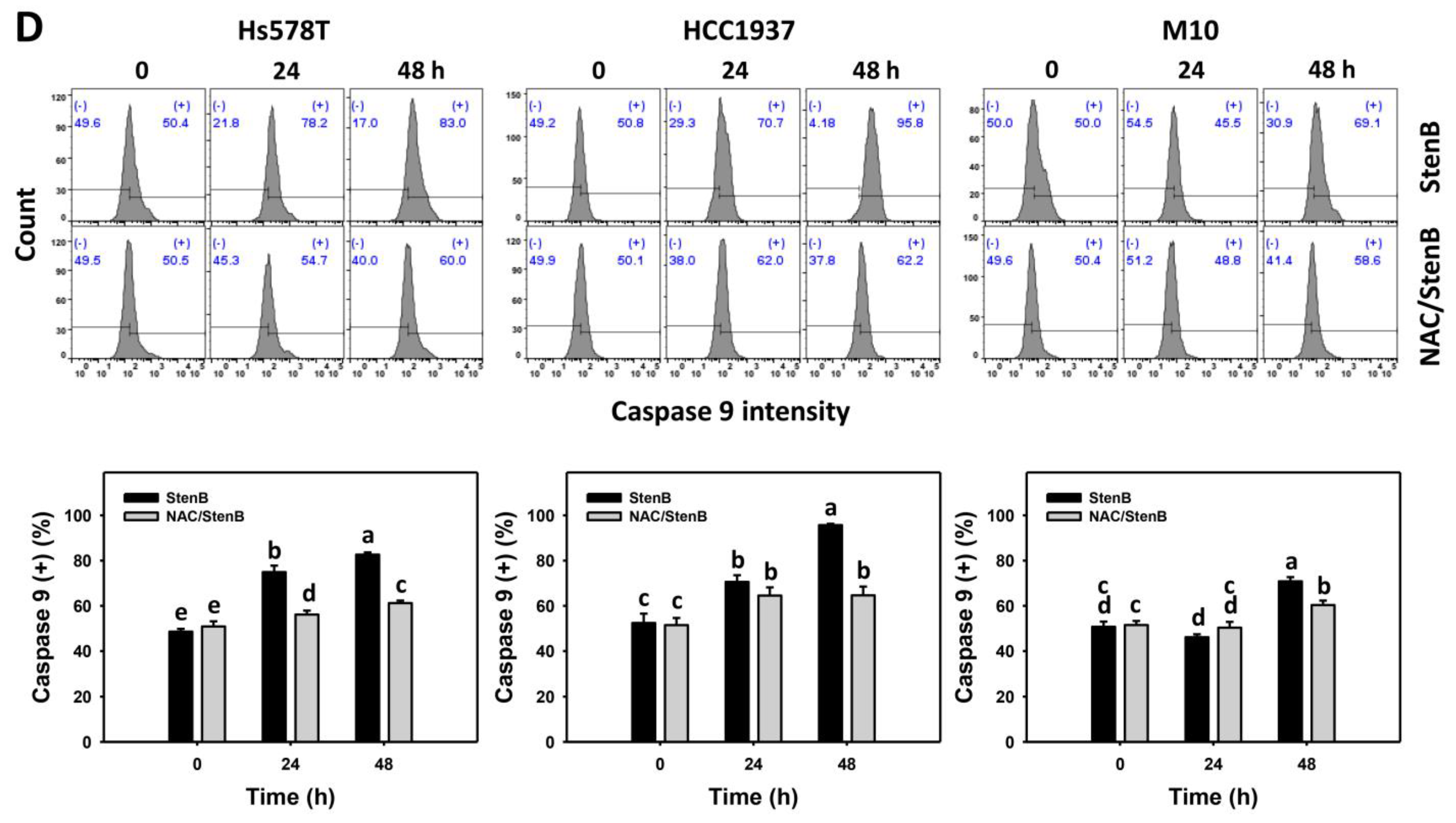
2.5. Caspase 8 and 9 Responses to StenB (TNBC vs. Normal Cells)
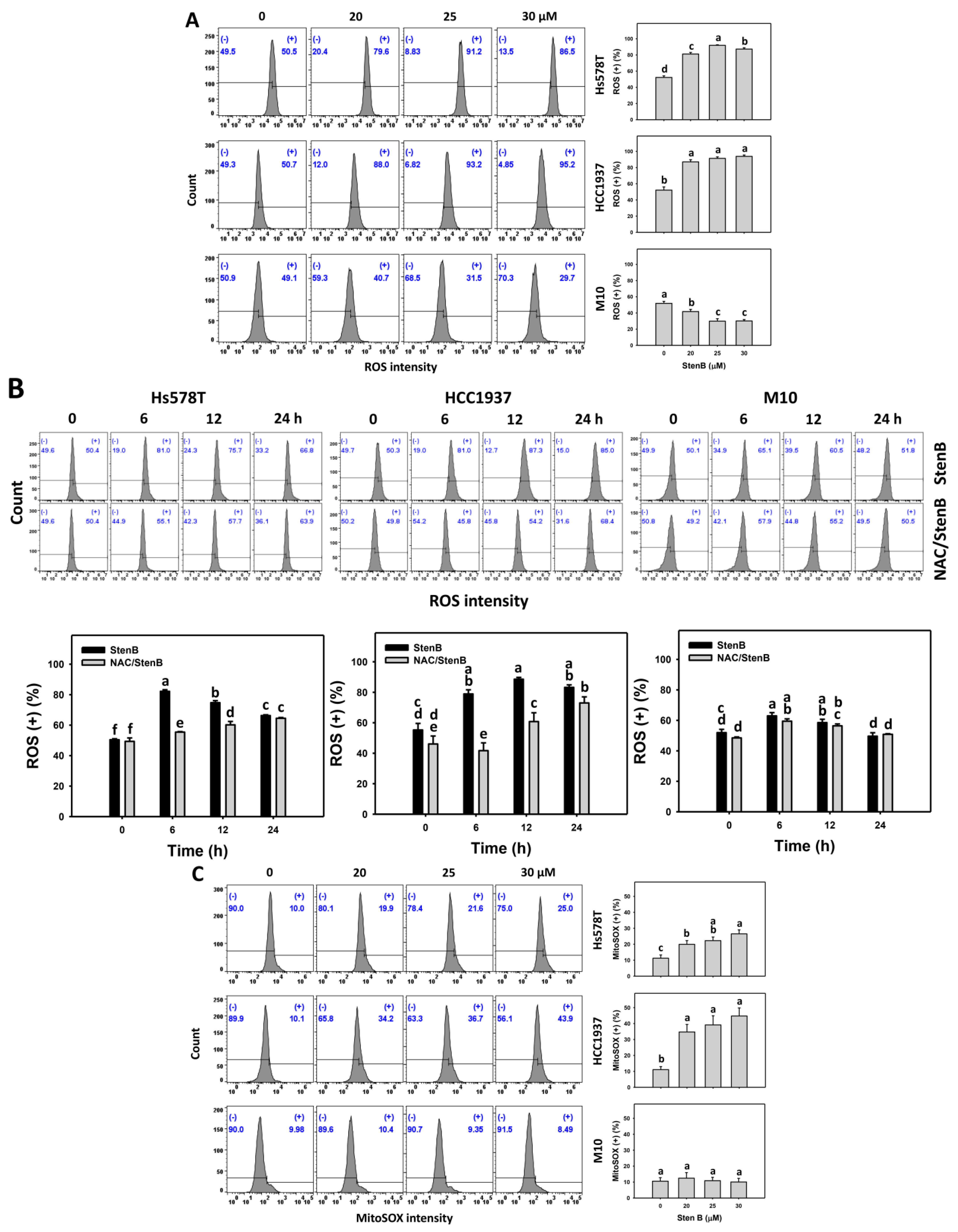
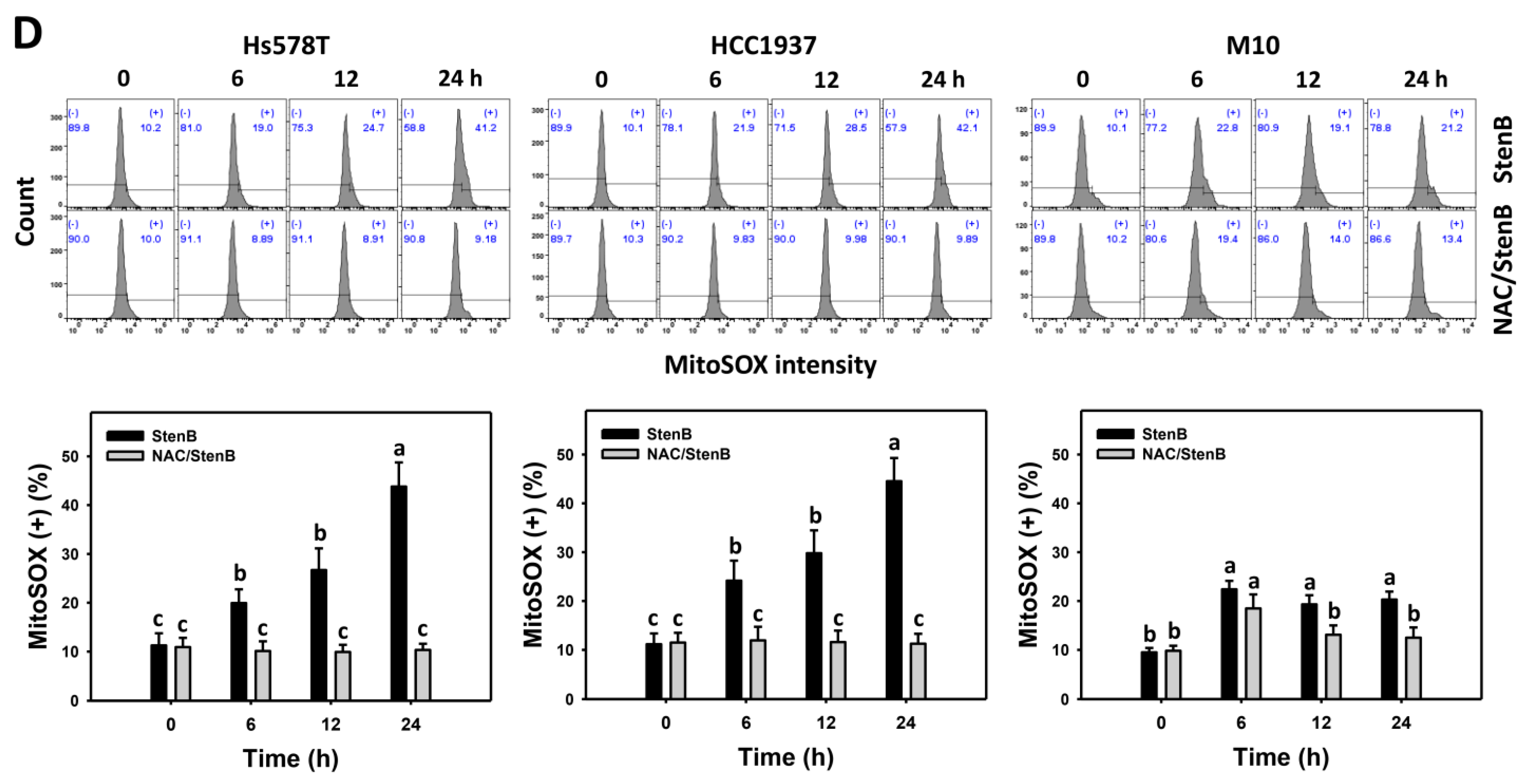
2.6. Reactive Oxygen Species (ROS) and Mitochondrial Superoxide (MitoSOX) Responses to StenB (TNBC vs. Normal Cells)
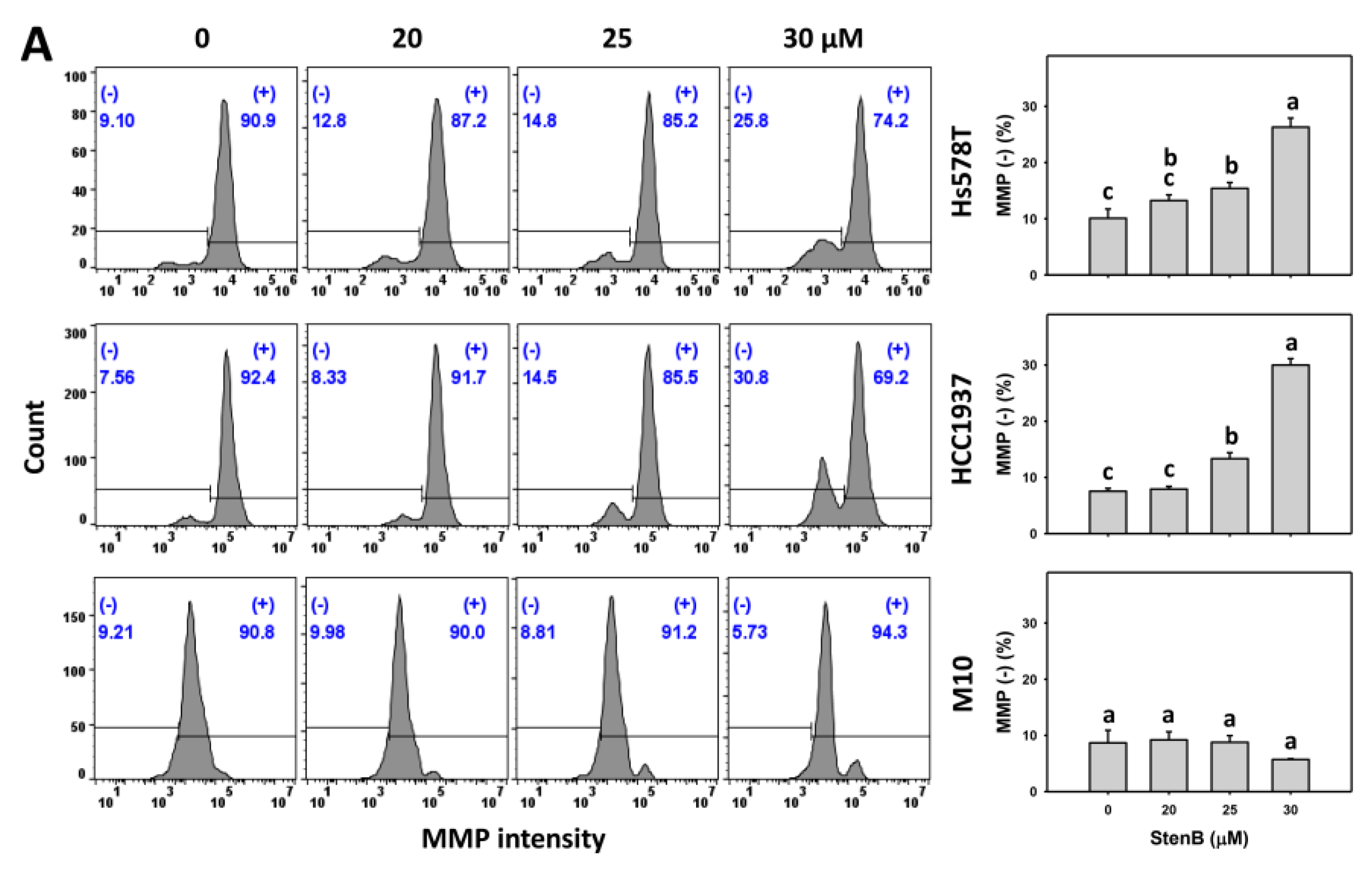
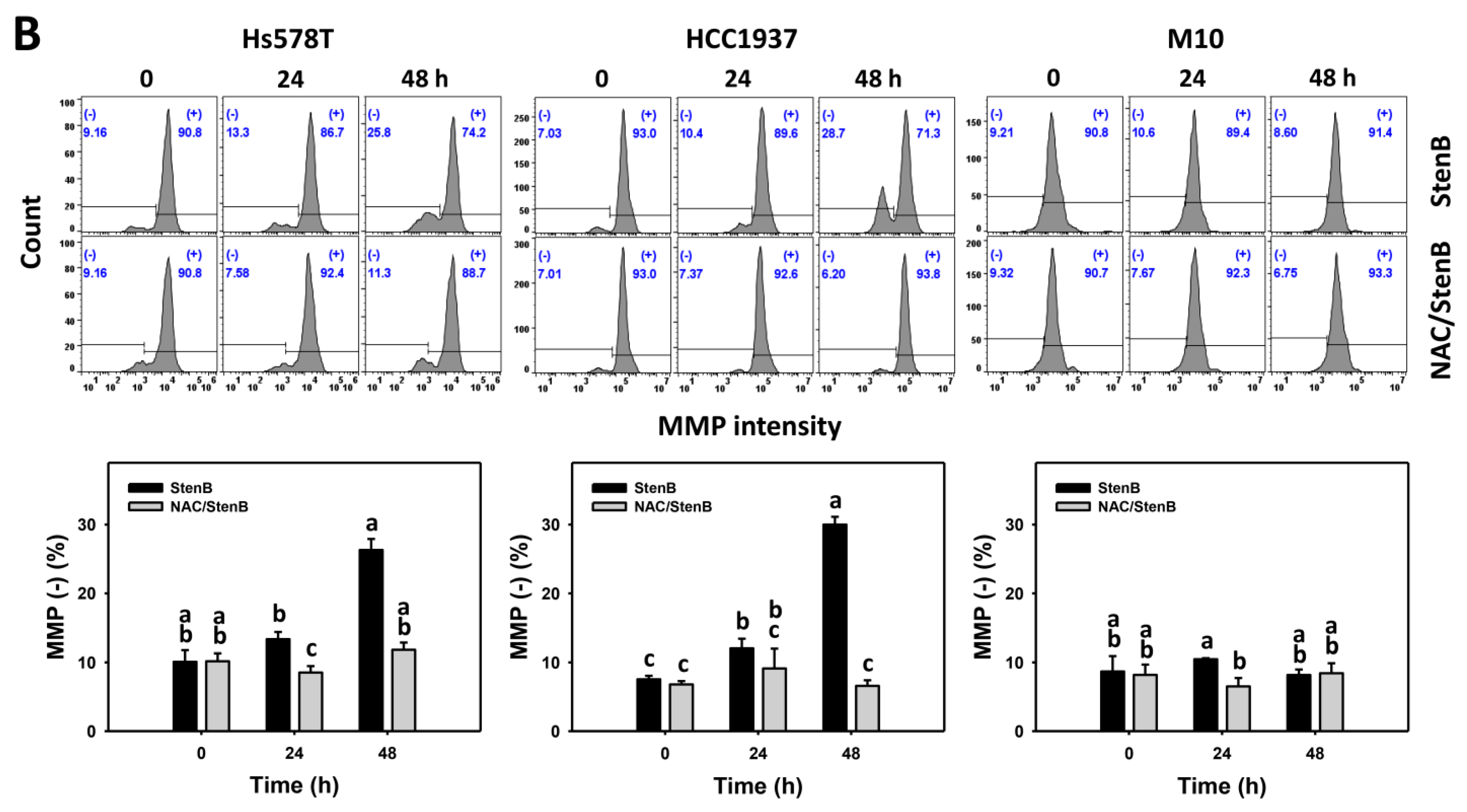
2.7. Mitochondrial Membrane Potential (MMP) Response to StenB (TNBC vs. Normal Cells)
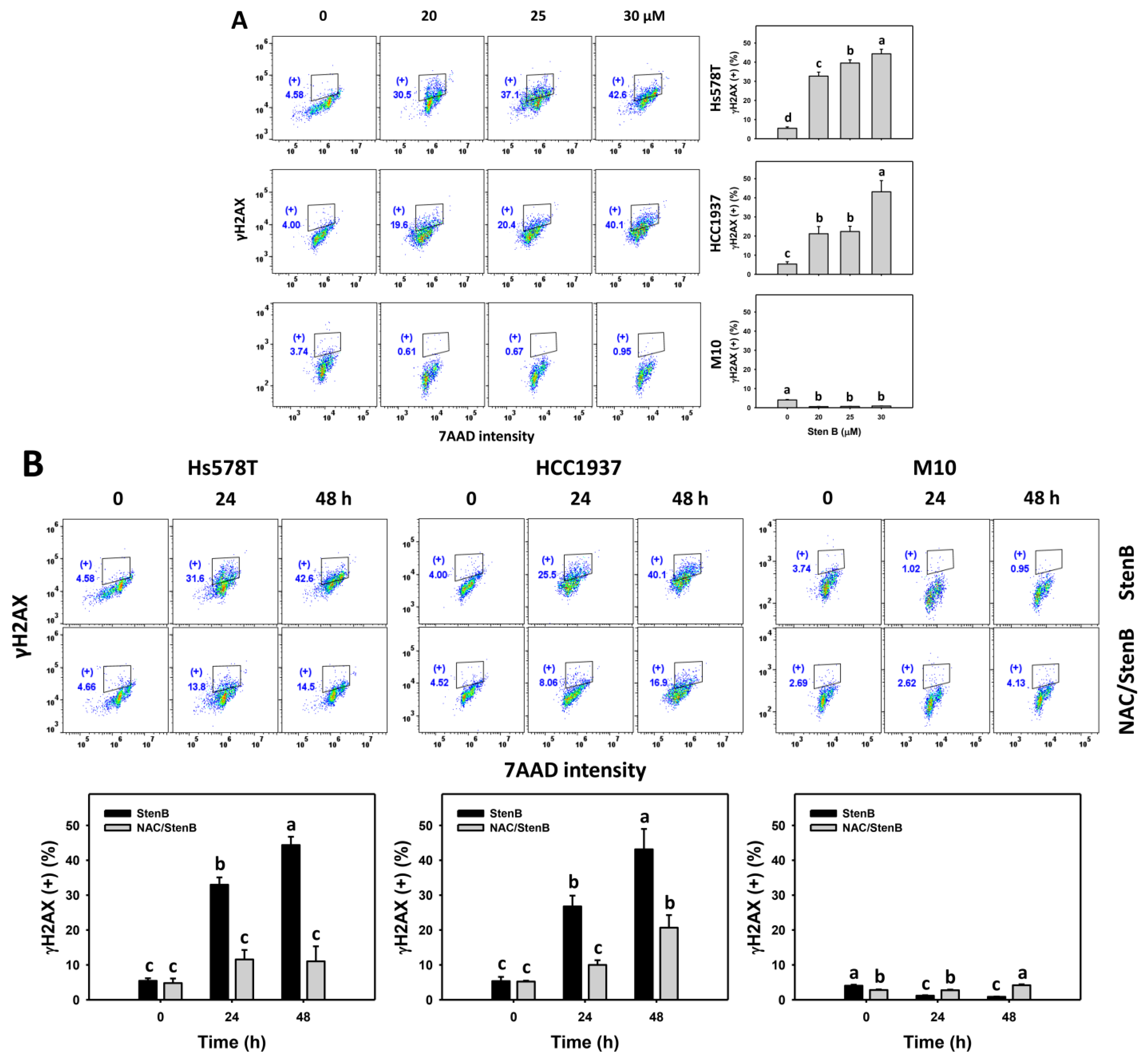
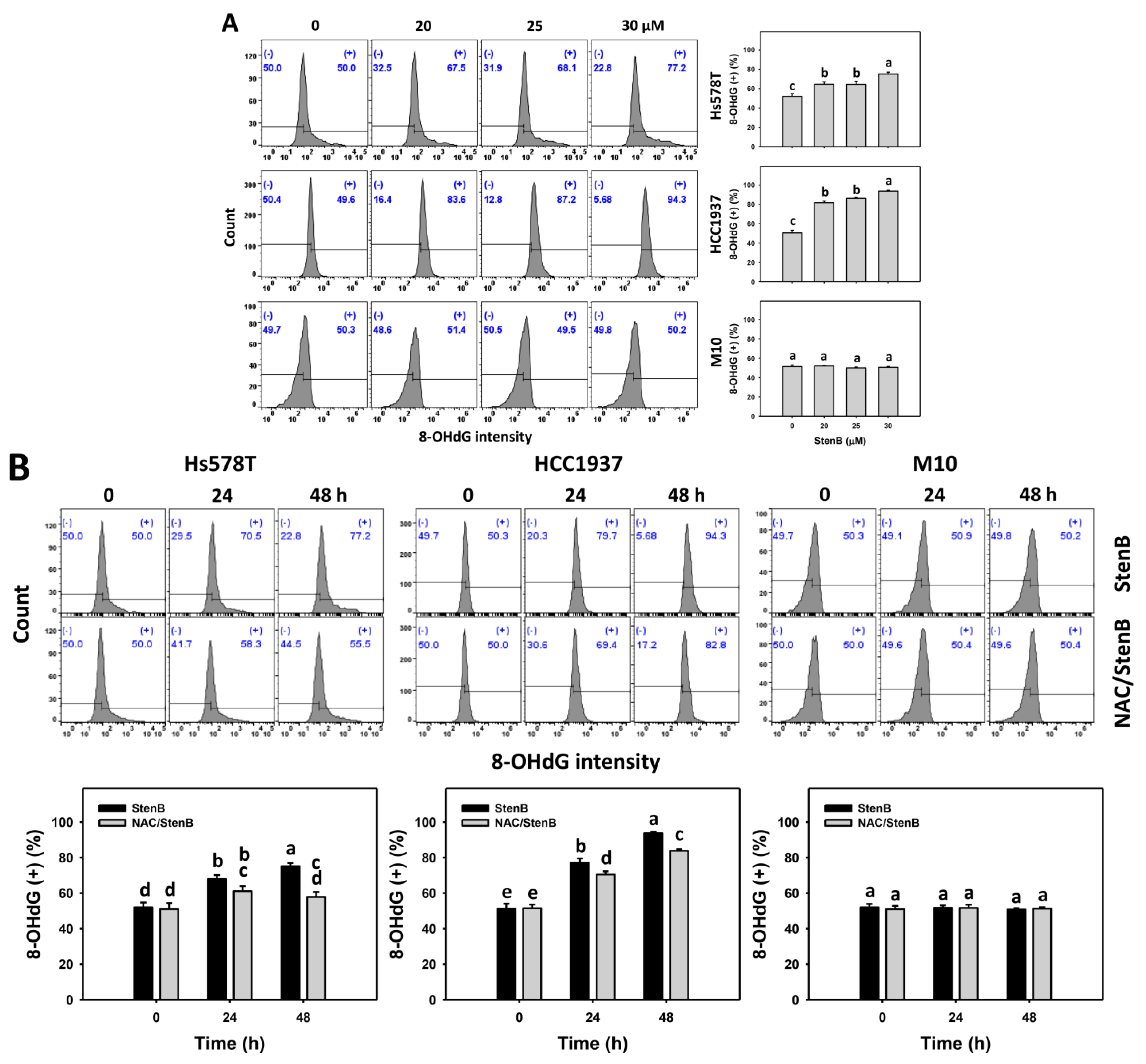
2.8. DNA Damage Responses to StenB (TNBC vs. Normal Cells)
3. Discussion
4. Materials and Methods
4.1. Preparation of StenB and Reagents
4.2. Cell Cultures and Viability
4.3. Cell Cycle
4.4. Apoptosis and Caspases 3, 8, and 9
4.5. Oxidative Stress
4.6. DNA Damages
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast cancer treatment: A review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- McCann, K.E.; Hurvitz, S.A.; McAndrew, N. Advances in targeted therapies for triple-negative breast cancer. Drugs 2019, 79, 1217–1230. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Castro, A.C.; Lin, N.U.; Polyak, K. Insights into molecular classifications of triple-negative breast cancer: Improving patient selection for treatment. Cancer Discov. 2019, 9, 176–198. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Chang, C.J.; Cheng, J.S. Survival, treatment regimens and medical costs of women newly diagnosed with metastatic triple-negative breast cancer. Sci. Rep. 2022, 12, 729. [Google Scholar] [CrossRef]
- Franzoi, M.A.; Agostinetto, E.; Perachino, M.; Del Mastro, L.; de Azambuja, E.; Vaz-Luis, I.; Partridge, A.H.; Lambertini, M. Evidence-based approaches for the management of side-effects of adjuvant endocrine therapy in patients with breast cancer. Lancet Oncol. 2021, 22, e303–e313. [Google Scholar] [CrossRef]
- Noor, A.; Md, N.; Halijah, I. Chemical constituents of essential oils of Boesenbergia armeniaca and B. stenophylla (Zingiberaceae) endemic to Borneo. Pak. J. Bot. 2018, 50, 1917–1922. [Google Scholar]
- Lam, N.F.; Ibrahim, H.; Sam, Y.Y.; Mohammad Zakaria, R.; Poulsen, A.D. Two new species of Boesenbergia (Zingiberaceae), from Sabah, Malaysia. PhytoKeys 2022, 211, 81–92. [Google Scholar] [CrossRef]
- Mohan, S.; Hobani, Y.H.; Shaheen, E.; Abou-Elhamd, A.S.; Abdelhaleem, A.; Alhazmi, H.A.; Abdelwahab, S.I. Ameliorative effect of Boesenbergin A, a chalcone isolated from Boesenbergia rotunda (Fingerroot) on oxidative stress and inflammation in ethanol-induced gastric ulcer in vivo. J. Ethnopharmacol. 2020, 261, 113104. [Google Scholar] [CrossRef]
- Abdelwahab, S.I.; Mohan, S.; Abdulla, M.A.; Sukari, M.A.; Abdul, A.B.; Taha, M.M.; Syam, S.; Ahmad, S.; Lee, K.H. The methanolic extract of Boesenbergia rotunda (L.) Mansf. and its major compound pinostrobin induces anti-ulcerogenic property in vivo: Possible involvement of indirect antioxidant action. J. Ethnopharmacol. 2011, 137, 963–970. [Google Scholar] [CrossRef]
- Ruttanapattanakul, J.; Wikan, N.; Okonogi, S.; Na Takuathung, M.; Buacheen, P.; Pitchakarn, P.; Potikanond, S.; Nimlamool, W. Boesenbergia rotunda extract accelerates human keratinocyte proliferation through activating ERK1/2 and PI3K/Akt kinases. Biomed. Pharmacother. 2021, 133, 111002. [Google Scholar] [CrossRef] [PubMed]
- Teethaisong, Y.; Pimchan, T.; Srisawat, R.; Hobbs, G.; Eumkeb, G. Boesenbergia rotunda (L.) Mansf. extract potentiates the antibacterial activity of some beta-lactams against beta-lactam-resistant staphylococci. J. Glob. Antimicrob. Resist. 2018, 12, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Zainin, N.; Lau, K.; Zakaria, M.; Son, R.; Razis, A.A.; Rukayadi, Y. Antibacterial activity of Boesenbergia rotunda (L.) Mansf. A. extract against Escherichia coli. Int. Food Res. J. 2013, 20, 3319. [Google Scholar]
- Kanjanasirirat, P.; Suksatu, A.; Manopwisedjaroen, S.; Munyoo, B.; Tuchinda, P.; Jearawuttanakul, K.; Seemakhan, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Rangkasenee, N.; et al. High-content screening of Thai medicinal plants reveals Boesenbergia rotunda extract and its component Panduratin A as anti-SARS-CoV-2 agents. Sci. Rep. 2020, 10, 19963. [Google Scholar] [CrossRef]
- Jirakiattikul, Y.; Rithichai, P.; Prachai, R.; Itharat, A. Elicitation enhancement of bioactive compound accumulation and antioxidant activity in shoot cultures of Boesenbergia rotunda L. Agric. Nat. Resour. 2021, 55, 456–463. [Google Scholar]
- Byahatti, S.D.; Thangadurai, D. Chemical constituents, antimicrobial potential and antioxidant efficacy of essential oil from Boesenbergia pulcherrima (Wall.) Kuntze. Plant. Arch. 2019, 19, 515–521. [Google Scholar]
- Jing, L.; Bakar, M.A.; Mohamed, M.; Rahmat, A. Effects of selected Boesenbergia species on the proliferation of several cancer cell lines. J. Pharmacol. Toxicol. 2011, 6, 272–282. [Google Scholar] [CrossRef]
- Pratama, M.R.F.; Praditapuspa, E.N.; Kesuma, D.; Poerwono, H.; Widiandani, T.; Siswodihardjo, S. Boesenbergia pandurata as an anti-breast cancer agent: Molecular docking and ADMET study. Lett. Drug. Des. Discov. 2022, 19, 606–626. [Google Scholar]
- Kirana, C.; Jones, G.P.; Record, I.R.; McIntosh, G.H. Anticancer properties of panduratin A isolated from Boesenbergia pandurata (Zingiberaceae). J. Nat. Med. 2007, 61, 131–137. [Google Scholar] [CrossRef]
- Win, N.N.; Awale, S.; Esumi, H.; Tezuka, Y.; Kadota, S. Bioactive secondary metabolites from Boesenbergia pandurata of Myanmar and their preferential cytotoxicity against human pancreatic cancer PANC-1 cell line in nutrient-deprived medium. J. Nat. Prod. 2007, 70, 1582–1587. [Google Scholar] [CrossRef]
- Break, M.K.B.; Chiang, M.; Wiart, C.; Chin, C.F.; Khoo, A.S.B.; Khoo, T.J. Cytotoxic activity of Boesenbergia rotunda extracts against nasopharyngeal carcinoma cells (HK1). Cardamonin, a Boesenbergia rotunda constituent, inhibits growth and migration of HK1 cells by inducing caspase-dependent apoptosis and G2/M-phase arrest. Nutr. Cancer 2021, 73, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Wongkaewkhiaw, S.; Wongrakpanich, A.; Krobthong, S.; Saengsawang, W.; Chairoungdua, A.; Boonmuen, N. Induction of apoptosis in human colorectal cancer cells by nanovesicles from fingerroot (Boesenbergia rotunda (L.) Mansf.). PLoS ONE 2022, 17, e0266044. [Google Scholar] [CrossRef] [PubMed]
- Primus, P.S.; Ismail, M.H.; Adnan, N.E.; Wu, C.H.Y.; Kao, C.L.; Choo, Y.M. Chemical constituents and anti-neuroblastoma activity from Boesenbergia stenophylla. Sains Malays. 2022, 51, 1075–1084. [Google Scholar] [CrossRef]
- Primus, P.S.; Ismail, M.H.; Adnan, N.E.; Wu, C.H.; Kao, C.L.; Choo, Y.M. Stenophyllols A-C, new compounds from Boesenbergia stenophylla. J. Asian Nat. Prod. Res. 2022, 24, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.b.; Jantan, I.b. The essential oils of Boesenbergia stenophylla RM Sm. as natural sources of methyl (E)-cinnamate. Flavour. Frag. J. 2003, 18, 485–486. [Google Scholar] [CrossRef]
- Mohammed, I.A.; Akhtar, M.N.; Biau, F.J.; Tor, Y.S.; Zareen, S.; Binti Shahabudin, S.; Binti Abd Hamid, H.; Ul Haq, Z.; Khalil, R.; Khalaf, R.M. Isolation of cardamonin and pinostrobin chalcone from the rhizomes of Boesenbergia rotunda (L.) Mansf. and their cytotoxic effects on H-29 and MDA-MB-231 cancer cell lines. Nat. Prod. J. 2019, 9, 341–348. [Google Scholar] [CrossRef]
- Halasa, M.; Luszczki, J.J.; Dmoszynska-Graniczka, M.; Baran, M.; Okon, E.; Stepulak, A.; Wawruszak, A. Antagonistic interaction between histone deacetylase inhibitor: Cambinol and cisplatin-an isobolographic analysis in breast cancer in vitro models. Int. J. Mol. Sci. 2021, 22, 8573. [Google Scholar] [CrossRef]
- Qi, L.; Luo, Q.; Zhang, Y.; Jia, F.; Zhao, Y.; Wang, F. Advances in toxicological research of the anticancer drug cisplatin. Chem. Res. Toxicol. 2019, 32, 1469–1486. [Google Scholar] [CrossRef]
- Azmanova, M.; Pitto-Barry, A. Oxidative stress in cancer therapy: Friend or enemy? Chembiochem 2022, 23, e202100641. [Google Scholar] [CrossRef]
- Raavi, V.; Perumal, V.; Paul, S.F. Potential application of gamma-H2AX as a biodosimetry tool for radiation triage. Mutat. Res. Rev. Mutat. Res. 2021, 787, 108350. [Google Scholar] [CrossRef]
- Banjarnahor, C.T.U.; Hardiany, N.S.; Wahjoepramono, E.J.; Hariyanto, A.D.; Sadikin, M. High concentration of gamma-H2AX correlates with a marker of apoptotic suppression and PI3K/Akt pathway upregulation in glioblastoma multiforme. Oncol. Lett. 2023, 25, 149. [Google Scholar] [CrossRef] [PubMed]
- Shekaftik, S.O.; Nasirzadeh, N. 8-Hydroxy-2′-deoxyguanosine (8-OHdG) as a biomarker of oxidative DNA damage induced by occupational exposure to nanomaterials: A systematic review. Nanotoxicology 2021, 15, 850–864. [Google Scholar] [CrossRef] [PubMed]
- AbuArrah, M.; Setianto, B.Y.; Faisal, A.; Sadewa, A.H. 8-Hydroxy-2-deoxyguanosine as oxidative DNA damage biomarker of medical ionizing radiation: A scoping review. J. Biomed. Phys. Eng. 2021, 11, 389. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.L.; Ho, H.Y.; Wu, Y.H.; Chiu, D.T. Glucose-6-phosphate dehydrogenase-deficient cells show an increased propensity for oxidant-induced senescence. Free. Radic. Biol. Med. 2004, 36, 580–591. [Google Scholar] [CrossRef]
- Takeda, S.; Hirao-Suzuki, M.; Shindo, M.; Aramaki, H. (–)-Xanthatin as a killer of human breast cancer MCF-7 mammosphere cells: A comparative study with salinomycin. Curr. Issues Mol. Biol. 2022, 44, 3849–3858. [Google Scholar] [CrossRef]
- Yamashita, N.; Taga, C.; Ozawa, M.; Kanno, Y.; Sanada, N.; Kizu, R. Camalexin, an indole phytoalexin, inhibits cell proliferation, migration, and mammosphere formation in breast cancer cells via the aryl hydrocarbon receptor. J. Nat. Med. 2022, 76, 110–118. [Google Scholar] [CrossRef]
- Bahadur Gurung, A.; Ajmal Ali, M.; Al-Hemaid, F.; El-Zaidy, M.; Lee, J. In silico analyses of major active constituents of fingerroot (Boesenbergia rotunda) unveils inhibitory activities against SARS-CoV-2 main protease enzyme. Saudi J. Biol. Sci. 2022, 29, 65–74. [Google Scholar] [CrossRef]
- Huang, C.H.; Yeh, J.M.; Chan, W.H. Hazardous impacts of silver nanoparticles on mouse oocyte maturation and fertilization and fetal development through induction of apoptotic processes. Environ. Toxicol. 2018, 33, 1039–1049. [Google Scholar] [CrossRef]
- Liu, Y.C.; Peng, B.R.; Hsu, K.C.; El-Shazly, M.; Shih, S.P.; Lin, T.E.; Kuo, F.W.; Chou, Y.C.; Lin, H.Y.; Lu, M.C. 13-Acetoxysarcocrassolide exhibits cytotoxic activity against oral cancer cells through the interruption of the Keap1/Nrf2/p62/SQSTM1 pathway: The need to move beyond classical concepts. Mar. Drugs 2020, 18, 382. [Google Scholar] [CrossRef]
- Shih, S.P.; Lu, M.C.; El-Shazly, M.; Lin, Y.H.; Chen, C.L.; Yu, S.S.F.; Liu, Y.C. The antileukemic and anti-prostatic effect of aeroplysinin-1 is mediated through ROS-induced apoptosis via NOX activation and inhibition of HIF-1a activity. Life 2022, 12, 687. [Google Scholar] [CrossRef]
- Yang, C.Y.; Lee, M.Y.; Chen, Y.L.; Shiau, J.P.; Tsai, Y.H.; Yang, C.N.; Chang, H.W.; Tseng, C.H. Synthesis and anticancer evaluation of 4-anilinoquinolinylchalcone derivatives. Int. J. Mol. Sci. 2023, 24, 6034. [Google Scholar] [CrossRef]
- Chen, C.Y.; Yen, C.Y.; Wang, H.R.; Yang, H.P.; Tang, J.Y.; Huang, H.W.; Hsu, S.H.; Chang, H.W. Tenuifolide B from Cinnamomum tenuifolium stem selectively inhibits proliferation of oral cancer cells via apoptosis, ROS generation, mitochondrial depolarization, and DNA damage. Toxins 2016, 8, 319. [Google Scholar] [CrossRef]
- Fan, H.C.; Hsieh, Y.C.; Li, L.H.; Chang, C.C.; Janouskova, K.; Ramani, M.V.; Subbaraju, G.V.; Cheng, K.T.; Chang, C.C. Dehydroxyhispolon methyl ether, a hispolon derivative, inhibits WNT/beta-catenin signaling to elicit human colorectal carcinoma cell apoptosis. Int. J. Mol. Sci. 2020, 21, 8839. [Google Scholar] [CrossRef] [PubMed]
- Rafat, A.; Dizaji Asl, K.; Mazloumi, Z.; Movassaghpour, A.A.; Talebi, M.; Shanehbandi, D.; Farahzadi, R.; Nejati, B.; Nozad Charoudeh, H. Telomerase inhibition on acute myeloid leukemia stem cell induced apoptosis with both intrinsic and extrinsic pathways. Life Sci. 2022, 295, 120402. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Chan, H.S.; Tsay, H.S.; Funayama, S.; Kuo, C.L.; Chung, J.G. Ethyl acetate fraction from methanol extraction of Vitis thunbergii var. taiwaniana induced G0 /G1 phase arrest via inhibition of cyclins D and E and induction of apoptosis through caspase-dependent and -independent pathways in human prostate carcinoma DU145 cells. Environ. Toxicol. 2018, 33, 41–51. [Google Scholar]
- Wu, C.F.; Lee, M.G.; El-Shazly, M.; Lai, K.H.; Ke, S.C.; Su, C.W.; Shih, S.P.; Sung, P.J.; Hong, M.C.; Wen, Z.H.; et al. Isoaaptamine induces T-47D cells apoptosis and autophagy via oxidative stress. Mar. Drugs 2018, 16, 18. [Google Scholar] [CrossRef] [PubMed]
- Shiau, J.P.; Chuang, Y.T.; Yang, K.H.; Chang, F.R.; Sheu, J.H.; Hou, M.F.; Jeng, J.H.; Tang, J.Y.; Chang, H.W. Brown algae-derived fucoidan exerts oxidative stress-dependent antiproliferation on oral cancer cells. Antioxidants 2022, 11, 841. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.H.; Lin, Y.S.; Wang, S.C.; Lee, M.Y.; Tang, J.Y.; Chang, F.R.; Chuang, Y.T.; Sheu, J.H.; Chang, H.W. Soft coral-derived dihydrosinularin exhibits antiproliferative effects associated with apoptosis and DNA damage in oral cancer cells. Pharmaceuticals 2021, 14, 994. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-Y.; Shiau, J.-P.; Tang, J.-Y.; Hou, M.-F.; Primus, P.S.; Kao, C.-L.; Choo, Y.-M.; Chang, H.-W. Boesenbergia stenophylla-Derived Stenophyllol B Exerts Antiproliferative and Oxidative Stress Responses in Triple-Negative Breast Cancer Cells with Few Side Effects in Normal Cells. Int. J. Mol. Sci. 2023, 24, 7751. https://doi.org/10.3390/ijms24097751
Lee M-Y, Shiau J-P, Tang J-Y, Hou M-F, Primus PS, Kao C-L, Choo Y-M, Chang H-W. Boesenbergia stenophylla-Derived Stenophyllol B Exerts Antiproliferative and Oxidative Stress Responses in Triple-Negative Breast Cancer Cells with Few Side Effects in Normal Cells. International Journal of Molecular Sciences. 2023; 24(9):7751. https://doi.org/10.3390/ijms24097751
Chicago/Turabian StyleLee, Min-Yu, Jun-Ping Shiau, Jen-Yang Tang, Ming-Feng Hou, Phoebe Sussana Primus, Chai-Lin Kao, Yeun-Mun Choo, and Hsueh-Wei Chang. 2023. "Boesenbergia stenophylla-Derived Stenophyllol B Exerts Antiproliferative and Oxidative Stress Responses in Triple-Negative Breast Cancer Cells with Few Side Effects in Normal Cells" International Journal of Molecular Sciences 24, no. 9: 7751. https://doi.org/10.3390/ijms24097751