
Chromosomal Characterization of Tripidium arundinaceum Revealed by Oligo-FISH

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Sequence Alignment Analysis between MCP Sequences and Sorghum Genome
2.2. Chromosome Painting Using MCPs in Sorghum
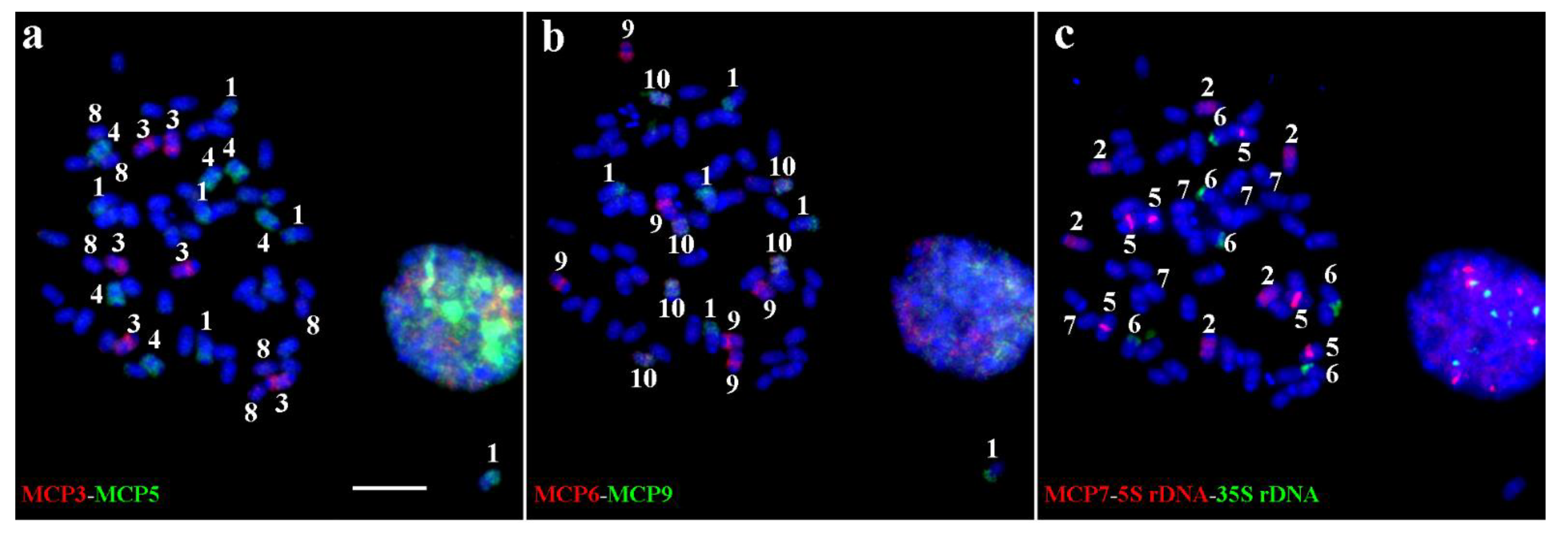
2.3. Chromosome Identification in T. arundinaceum Using MCPs, 5S rDNA, and 35S rDNA Probes
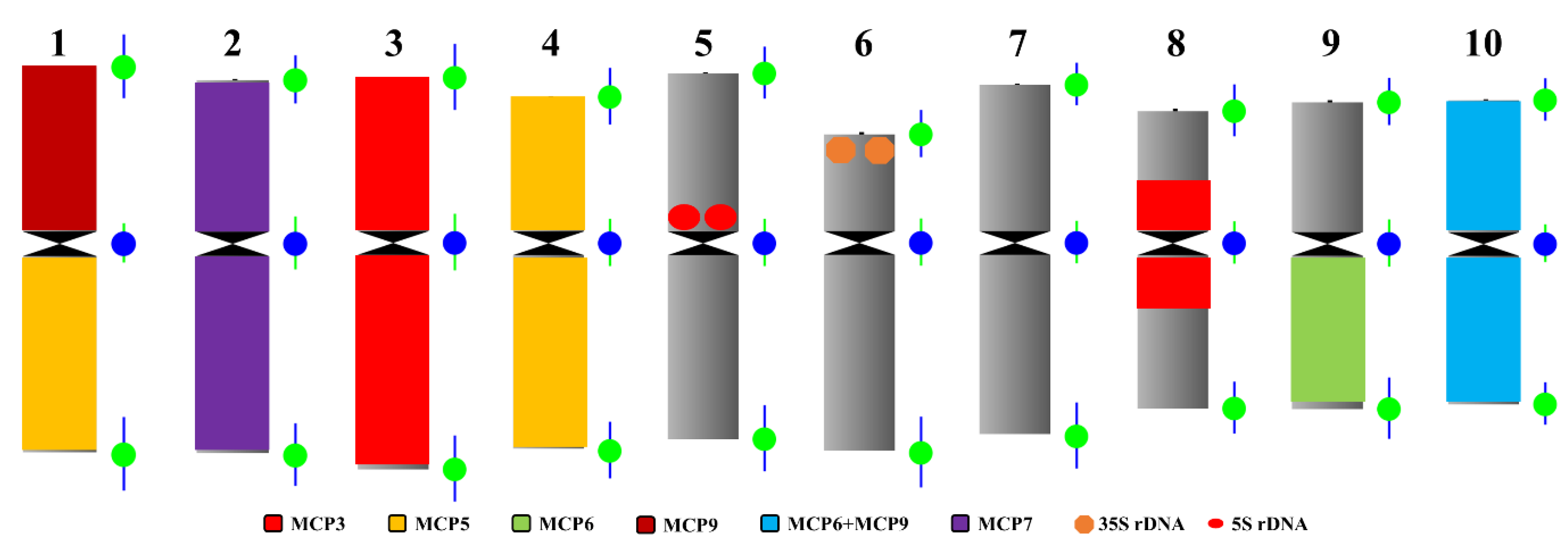
2.4. Standard Karyotype Analysis of T. arundinaceum Based on Sequential Oligo-FISH
3. Discussion
4. Materials and Methods
4.1. Plant Material and the Preparation of Metaphase Plates
4.2. Sequence Alignment and Analysis
4.3. Oligo-FISH and Karyotype Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roach, B.T. Nobilisation of sugarcane. Proc. Int. Soc. Sugar Cane Technol. 1972, 14, 206–216. [Google Scholar]
- Anzoua, K.G.; Yamada, T.; Henry, R.J. Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Lloyd Evans, D.; Joshi, S.V.; Wang, J. Whole chloroplast genome and gene locus phylogenies reveal the taxonomic placement and relationship of Tripidium (Panicoideae: Andropogoneae) to sugarcane. BMC Evol. Biol. 2019, 19, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachakkil, B.; Terajima, Y.; Ohmido, N.; Ebina, M.; Irei, S.; Hayashi, H.; Takagi, H. Cytogenetic and agronomic characterization of intergeneric hybrids between Saccharum spp. hybrid and Erianthus arundinaceus. Sci. Rep. 2019, 9, 1748. [Google Scholar] [CrossRef] [Green Version]
- Piperidis, N.; Chen, J.W.; Deng, H.H.; Wang, L.P.; Jackson, P.; Piperidis, G. GISH characterization of Erianthus arundinaceus chromosomes in three generations of sugarcane intergeneric hybrids. Genome 2010, 53, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zeng, K.; Chen, K.; Wu, J.; Wang, Q.; Li, X.; Deng, Z.; Huang, Y.; Huang, F.; Chen, R.; et al. Chromosome transmission in BC4 progenies of intergeneric hybrids between Saccharum spp. and Erianthus arundinaceus (Retz.) Jeswiet. Sci. Rep. 2019, 9, 2528. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J. Fluorescence in situ hybridization in plants: Recent developments and future applications. Chromosome Res. 2019, 27, 153–165. [Google Scholar] [CrossRef]
- Jiang, J.; Gill, B.S. Nonisotopic in situ hybridization and plant genome mapping: The first 10 years. Genome 1994, 37, 717–725. [Google Scholar] [CrossRef]
- Singh, R.S.; Jiang, J.; Gill, B.S. Current status and the future of fluorescence in situ hybridization (FISH) in plant genome research. Genome 2006, 49, 1057–1068. [Google Scholar]
- Zhang, T.; Liu, G.; Zhao, H.; Braz, G.T.; Jiang, J. Chorus2: Design of genome-scale oligonucleotide-based probes for fluorescence in situ hybridization. Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef]
- Liu, X.; Sun, S.; Wu, Y.; Zhou, Y.; Gu, S.; Yu, H.; Yi, C.; Gu, M.; Jiang, J.; Liu, B.; et al. Dual-color oligo-FISH can reveal chromosomal variations and evolution in Oryza species. Plant J. 2020, 101, 112–121. [Google Scholar] [CrossRef]
- Song, X.; Song, R.; Zhou, J.; Yan, W.; Zhang, T.; Sun, H.; Xiao, J.; Wu, Y.; Xi, M.; Lou, Q.; et al. Development and application of oligonucleotide-based chromosome painting for chromosome 4D of Triticum aestivum L. Chromosome Res. 2020, 28, 171–182. [Google Scholar] [CrossRef]
- He, L.; Zhao, H.; He, J.; Yang, Z.; Jiang, J. Extraordinarily conserved chromosomal synteny of Citrus species revealed by chromosome: Pecific painting. Plant J. 2020, 103, 2225–2235. [Google Scholar] [CrossRef]
- Albert, P.S.; Zhang, T.; Semrau, K.; Rouillard, J.M.; Kao, Y.H.; Wang, C.J.R.; Danilova, T.V.; Jiang, J.; Birchler, J.A. Whole-chromosome paints in maize reveal rearrangements, nuclear domains, and chromosomal relationships. Proc. Natl. Acad. Sci. USA 2019, 116, 1679–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braz, G.T.; He, L.; Zhao, H.; Zhang, T.; Jiang, J. Comparative Oligo-FISH Mapping: An Efficient and Powerful Methodology To Reveal Karyotypic and Chromosomal Evolution. Genetics 2017, 208, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, H.; Zhang, T.; Wu, Y.; Zhang, W.; Zhang, P.; Xi, M.; Jiang, J. An extraordinarily stable karyotype of the woody Populus species revealed by chromosome painting. Plant J. 2020, 101, 253–264. [Google Scholar] [CrossRef]
- Shin-Ichi, T.; Masumi, E.; Makoto, K.; Wataru, T.; Berthold, H. Complete Chloroplast Genomes of Erianthus arundinaceus and Miscanthus sinensis: Comparative Genomics and Evolution of the Saccharum Complex. PLoS ONE 2017, 12, e0169992. [Google Scholar]
- Levan, A. Nomenclature for centromeric position on chromosomes. Heriditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Zhou, H.C.; Pellerin, R.J.; Waminal, N.E.; Yang, T.-J.; Kim, H.H. Pre-labelled oligo probe-FISH karyotype analyses of four Araliaceae species using rDNA and telomeric repeat. Genes Genom. 2019, 41, 839–847. [Google Scholar] [CrossRef]
- Do Vale Martins, L.; de Oliveira Bustamante, F.; da Silva Oliveira, A.R.; da Costa, A.F.; de Lima Feitoza, L.; Liang, Q.; Zhao, H.; Benko-Iseppon, A.M.; Muñoz-Amatriaín, M.; Pedrosa-Harand, A.; et al. BAC- and oligo-FISH mapping reveals chromosome evolution among Vigna angularis, V. unguiculata, and Phaseolus vulgaris. Chromosoma 2021, 1–15. [Google Scholar] [CrossRef]
- Guangrui, D.; Jiao, S.; Qing, Z.; Jianping, W.; Qingyi, Y.; Ray, M.; Kai, W.; Jisen, Z. Development and Applications of Chromosome-Specific Cytogenetic BAC-FISH Probes in S. spontaneum. Front. Plant Sci. 2018, 9, 218. [Google Scholar]
- Thumjamras, S.; Iamtham, S.; Prammanee, S.; de Jong, H. Meiotic analysis and FISH with rDNA and rice BAC probes of the Thai KPS 01-01-25 sugarcane cultivar. Plant Syst. Evol. 2016, 302, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Huang, Y.; Luo, L.; Li, X.; Wu, J.; Chen, R.; Zhang, M.; Deng, Z. An improved suppression subtractive hybridization technique to develop species-specific repetitive sequences from Erianthus arundinaceus (Saccharum complex). BMC Plant Biol. 2018, 18, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-Specific Painting in Cucumis Species Using Bulked Oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swigonova, Z. Close Split of Sorghum and Maize Genome Progenitors. Genome Res. 2004, 14, 1916–1923. [Google Scholar] [CrossRef] [Green Version]
- Braz, G.T.; do Vale Martins, L.; Zhang, T.; Albert, P.S.; Birchler, J.A.; Jiang, J. A universal chromosome identification system for maize and wild Zea species. Chromosome Res. 2020, 28, 183–194. [Google Scholar] [CrossRef]
- Besse, P.; Taylor, G.; Carroll, B.; Berding, N.; Burner, D.; McIntyre, C.L. Assessing genetic diversity in a sugarcane germplasm collection using an automated AFLP analysis. Genetica 1998, 104, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Zhang, Z.; Yan, T.; Lin, Q.; Wang, Y.; Huang, W.; Huang, Y.; Li, Z.; Yu, Q.; Wang, J.; et al. Comprehensively Characterizing the Cytological Features of Saccharum spontaneum by the Development of a Complete Set of Chromosome-Specific Oligo Probes. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Jiayun, W.; Yongji, H.; Yanquan, L.; Cheng, F.; Shaomou, L.; Zuhu, D.; Qiwei, L.; Zhongxing, H.; Rukai, C.; Muqing, Z. Unexpected Inheritance Pattern of Erianthus arundinaceus Chromosomes in the Intergeneric Progeny between Saccharum spp. and Erianthus arundinaceus. PLoS ONE 2014, 9, e110390. [Google Scholar]
- Braz, G.T.; Yu, F.; do Vale Martins, L.; Jiang, J. Fluorescent In Situ Hybridization Using Oligonucleotide-Based Probes. Methods Mol. Bio. 2020, 2148, 71–83. [Google Scholar]
- Chengjie, C.; Hao, C.; Yi, Z.; Hannah, R.T.; Margaret, H.F.; Yehua, H.; Rui, X. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hao, Z. RIdeogram: Drawing SVG graphics to visualize and map genome-wide data on the idiograms. PeerJ Comput. Sci. 2020, 6, e251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Ding, W.; Zhang, M.; Han, J.; Jing, Y.; Yao, W.; Hasterok, R.; Wang, Z.; Wang, K. The formation and evolution of centromeric satellite repeats in Saccharum species. Plant J. 2021, 106, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Kirov, I.; Khrustaleva, L.; Van Laere, K.; Soloviev, A.; Meeus, S.; Romanov, D.; Fesenko, I. DRAWID: User-friendly java software for chromosome measurements and idiogram drawing. Comp. Cytogenet. 2017, 11, 747–757. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Oligo | Sorghum1 | Sorghum2 | Sorghum3 | Sorghum4 | Sorghum5 | Sorghum6 | Sorghum7 | Sorghum8 | Sorghum9 | Sorghum10 |
|---|---|---|---|---|---|---|---|---|---|---|
| MCP1 | 27,169 | 746 | 892 | 820 | 657 | 689 | 4812 | 2772 | 700 | 791 |
| MCP2 | 701 | 9016 | 564 | 521 | 2613 | 11,784 | 405 | 446 | 525 | 524 |
| MCP3 | 711 | 546 | 20,574 | 550 | 495 | 415 | 463 | 3761 | 435 | 513 |
| MCP4 | 836 | 531 | 600 | 9900 | 5120 | 449 | 4999 | 564 | 484 | 543 |
| MCP5 | 7236 | 539 | 552 | 15,991 | 382 | 468 | 411 | 490 | 530 | 2548 |
| MCP6 | 625 | 416 | 507 | 404 | 424 | 322 | 1390 | 659 | 10,918 | 6910 |
| MCP7 | 488 | 17,672 | 421 | 422 | 316 | 316 | 387 | 347 | 367 | 415 |
| MCP8 | 466 | 402 | 11,297 | 420 | 294 | 282 | 313 | 372 | 5725 | 419 |
| MCP9 | 8544 | 434 | 412 | 411 | 277 | 304 | 296 | 300 | 329 | 9614 |
| MCP10 | 648 | 471 | 501 | 479 | 466 | 9307 | 2888 | 3983 | 1136 | 481 |
| Chromosome Number | Long Arm (μm) | Short Arm (μm) | Chromosome Length (μm) | Arm Ratio |
|---|---|---|---|---|
| Chr.1 | 2.02 (±0.19) | 1.56 (±0.10) | 3.58 (±0.19) | 1.30 (±0.18) |
| Chr.2 | 2.03 (±0.06) | 1.54 (±0.10) | 3.57 (±0.11) | 1.32 (±0.11) |
| Chr.3 | 2.08 (±0.21) | 1.49 (±0.21) | 3.57 (±0.30) | 1.42 (±0.24) |
| Chr.4 | 1.84 (±0.16) | 1.29 (±0.15) | 3.13 (±0.22) | 1.45 (±0.21) |
| Chr.5 | 1.76 (±0.21) | 1.49 (±0.13) | 3.25 (±0.26) | 1.19 (±0.17) |
| Chr.6 | 1.80 (±0.90) | 0.90 (±0.10) | 2.69 (±0.22) | 2.03 (±0.37) |
| Chr.7 | 1.70 (±0.19) | 1.39 (±0.09) | 3.10 (±0.24) | 1.22 (±0.12) |
| Chr.8 | 1.40 (±0.13) | 1.10 (±0.14) | 2.50 (±0.23) | 1.29 (±0.15) |
| Chr.9 | 1.42 (±0.16) | 1.19 (±0.11) | 2.61 (±0.20) | 1.20 (±0.18) |
| Chr.10 | 1.38 (±0.08) | 1.21 (±0.09) | 2.59 (±0.12) | 1.14 (±0.10) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Chai, J.; Li, X.; Yu, Z.; Yang, R.; Ding, X.; Wang, Q.; Wu, J.; Yang, X.; Deng, Z. Chromosomal Characterization of Tripidium arundinaceum Revealed by Oligo-FISH. Int. J. Mol. Sci. 2021, 22, 8539. https://doi.org/10.3390/ijms22168539
Yu F, Chai J, Li X, Yu Z, Yang R, Ding X, Wang Q, Wu J, Yang X, Deng Z. Chromosomal Characterization of Tripidium arundinaceum Revealed by Oligo-FISH. International Journal of Molecular Sciences. 2021; 22(16):8539. https://doi.org/10.3390/ijms22168539
Chicago/Turabian StyleYu, Fan, Jin Chai, Xueting Li, Zehuai Yu, Ruiting Yang, Xueer Ding, Qiusong Wang, Jiayun Wu, Xiping Yang, and Zuhu Deng. 2021. "Chromosomal Characterization of Tripidium arundinaceum Revealed by Oligo-FISH" International Journal of Molecular Sciences 22, no. 16: 8539. https://doi.org/10.3390/ijms22168539