Cuscuta Species Identification Based on the Morphology of Reproductive Organs and Complete Chloroplast Genome Sequences



Abstract
:1. Introduction
2. Results and Discussion
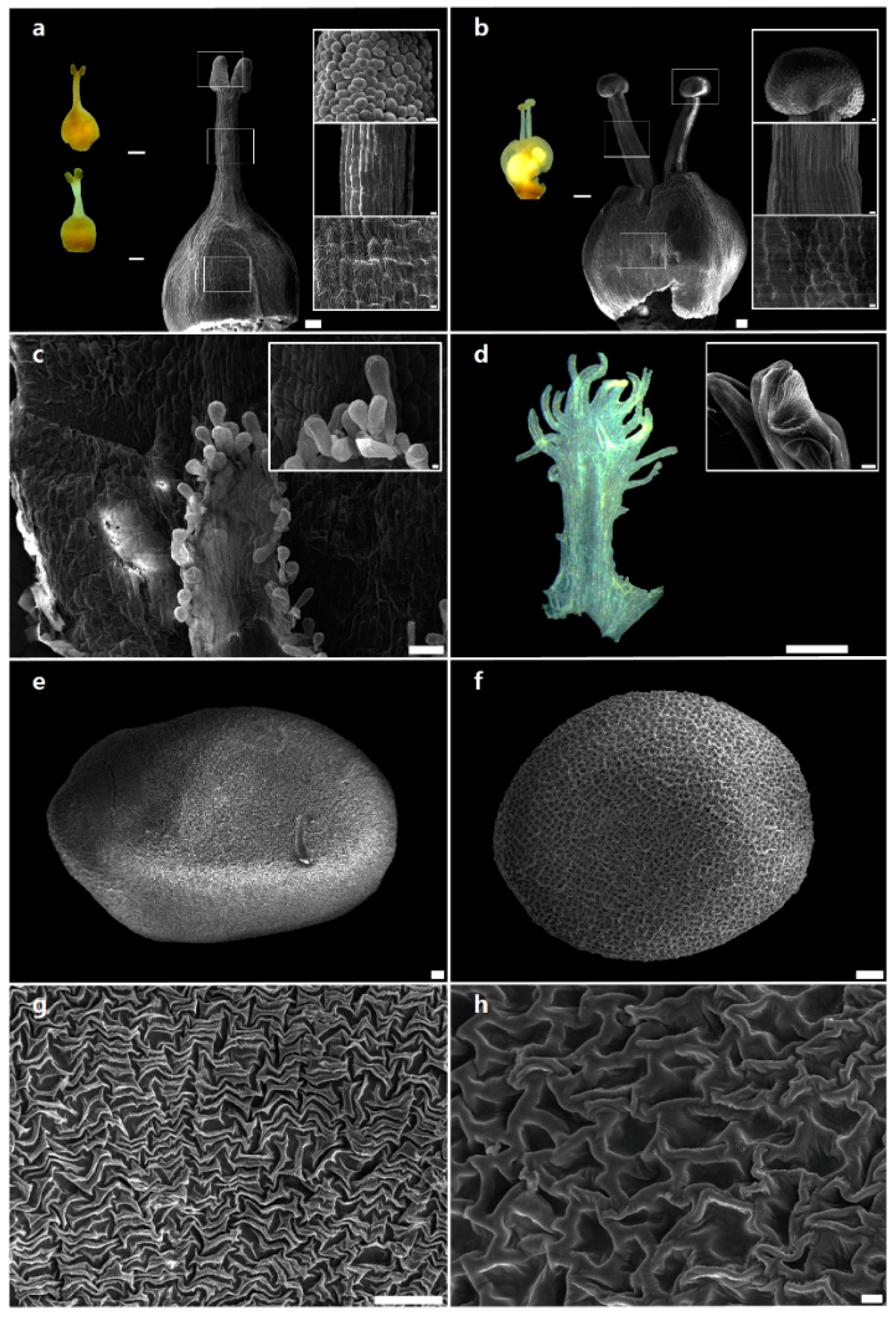
2.1. Morphological Characteristics of Reproductive Organs
2.2. Chloroplast Genome Features of C. japonica and C. chinensis
2.3. Dynamic Chloroplast Genome Structure of Cuscuta
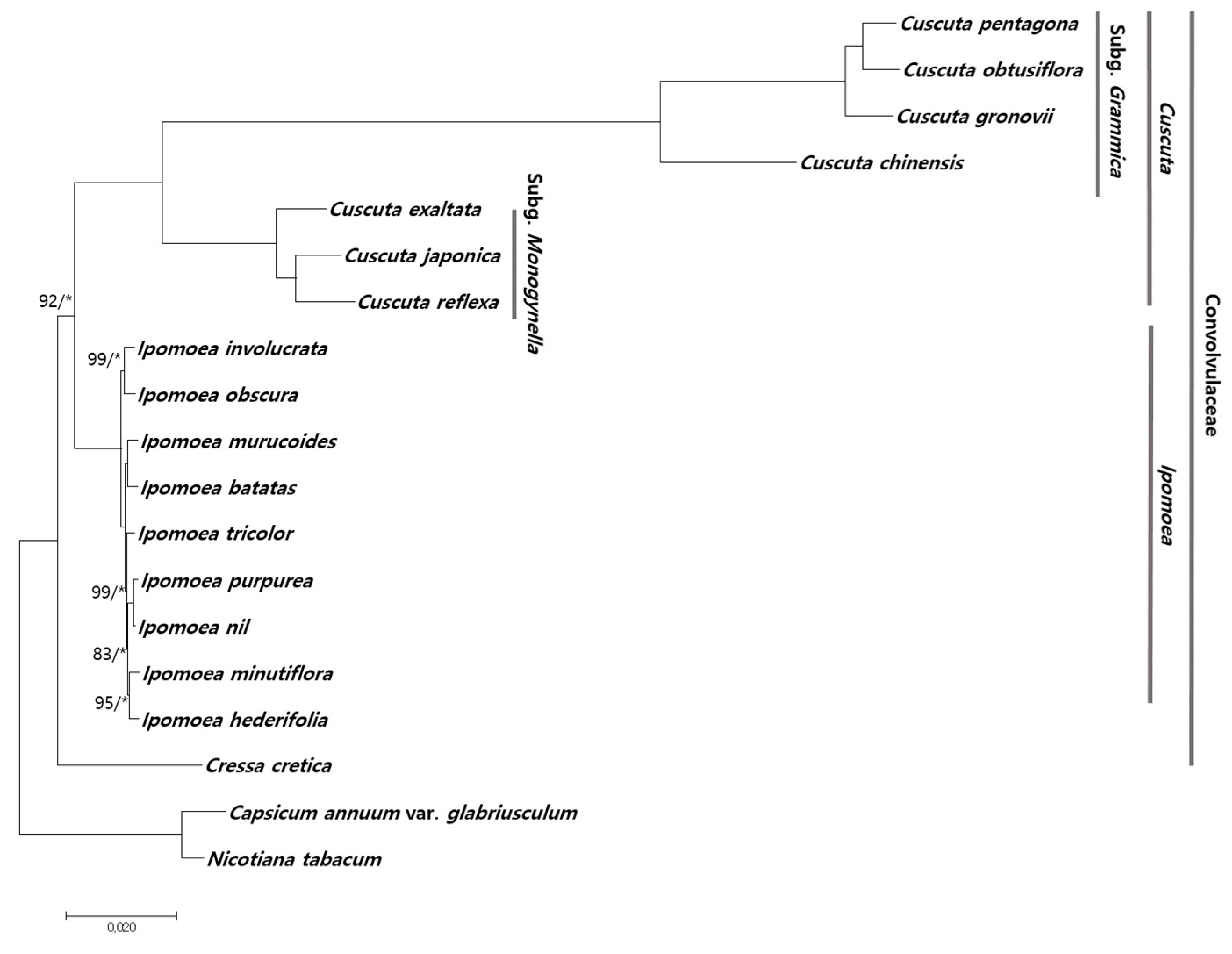
2.4. Phylogenetic Relationship among Cuscuta Species
3. Materials and Methods
3.1. Plant Material
3.2. Microscopic Analysis
3.3. Genome Sequencing and Assembly
3.4. Genome Annotation and Comparative Analysis
3.5. Repeat Analysis
3.6. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Costea, M.; García, M.A.; Stefanović, S. A phylogenetically based infrageneric classification of the parasitic plant genus Cuscuta (dodders, Convolvulaceae). Syst. Bot. 2015, 40, 269–285. [Google Scholar] [CrossRef]
- Fang, R.-C.; Musselman, L.J.; Plitmann, U. Cuscuta Linnaeus. In Flora of China, Gentianaceae Through Boraginaceae; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1995; Volume 16, pp. 322–325. [Google Scholar]
- Dawson, J.H.; Musselman, L.J.; Wolswinkel, P.; Dörr, I. Biology and control of Cuscuta. Rev. Weed Sci. 1994, 6, 265–317. [Google Scholar]
- Costea, M.; Tardif, F.J. The biology of Canadian weeds. 133. Cuscuta campestris Yuncker, C. gronovii Willd. ex Schult., C. umbrosa Beyr. ex Hook., C. epithymum (L.) L. and C. epilinum Weihe. Can. J. Plant Sci. 2006, 86, 293–316. [Google Scholar] [CrossRef]
- Hwang, S.; Kil, J.; Lee, C.-W.; Kim, Y. Distribution and host plants of parasitic weed Cuscuta pentagona Engelm. Hangug Jawon Sigmul Haghoeji 2013, 26, 289–302. [Google Scholar] [CrossRef]
- Costea, M.; Stefanović, S. Cuscuta jepsonii (Convolvulaceae): An invasive weed or an extinct endemic? Am. J. Bot. 2009, 96, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Chung, Y.; Oh, S.M. Taxonomic evaluation of selected Cuscuta species (Cuscutaceae) based on seed morphology. Kor. J. Weed Sci. 2000, 20, 255–263. [Google Scholar]
- Welsh, M.; Stefanović, S.; Costea, M. Pollen evolution and its taxonomic significance in Cuscuta (dodders, Convolvulaceae). Plant Syst. Evol. 2010, 285, 83–101. [Google Scholar] [CrossRef]
- Wright, M.A.; Welsh, M.; Costea, M. Diversity and evolution of the gynoecium in Cuscuta (dodders, Convolvulaceae) in relation to their reproductive biology: Two styles are better than one. Plant Syst. Evol. 2011, 296, 51–76. [Google Scholar] [CrossRef]
- Riviere, S.; Clayson, C.; Dockstader, K.; Wright, M.A.; Costea, M. To attract or to repel? Diversity, evolution and role of the “most peculiar organ” in the Cuscuta flower (dodder, Convolvulaceae)—the infrastaminal scales. Plant Syst. Evol. 2013, 299, 529–552. [Google Scholar] [CrossRef]
- Jansen, R.K.; Ruhlman, T.A. Plastid genomes of seed plants. In Genomics of Chloroplasts and Mitochondria; Springer: Dordrecht, The Netherlands, 2012; pp. 103–126. [Google Scholar]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H.; Depamphilis, C.W.; Leebens-Mack, J.; Muller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.; Yang, S.; Kim, W.J.; Noh, P.; Lee, H.O.; Moon, B.C. The complete chloroplast genomes of six Ipomoea species and indel marker development for the discrimination of authentic Pharbitidis Semen (seeds of I. nil or I. purpurea). Front. Plant Sci. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; de Pamphilis, C.W.; Muller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Frailey, D.C.; Chaluvadi, S.R.; Vaughn, J.N.; Coatney, C.G.; Bennetzen, J.L. Gene loss and genome rearrangement in the plastids of five hemiparasites in the family Orobanchaceae. BMC Plant Biol. 2018, 18, 30. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, J.G.; Chen, X.L.; Cui, Y.X.; Xu, Z.C.; Li, Y.H.; Song, J.Y.; Duan, B.Z.; Yao, H. Gene losses and partial deletion of small single-copy regions of the chloroplast genomes of two hemiparasitic Taxillus species. Sci. Rep. 2017, 7, 12834. [Google Scholar] [CrossRef] [PubMed]
- Funk, H.T.; Berg, S.; Krupinska, K.; Maier, U.G.; Krause, K. Complete DNA sequences of the plastid genomes of two parasitic flowering plant species, Cuscuta reflexa and Cuscuta gronovii. BMC Plant Biol. 2007, 7, 45. [Google Scholar] [CrossRef]
- McNeal, J.R.; Kuehl, J.V.; Boore, J.L.; de Pamphilis, C.W. Complete plastid genome sequences suggest strong selection for retention of photosynthetic genes in the parasitic plant genus Cuscuta. BMC Plant Biol. 2007, 7, 57. [Google Scholar] [CrossRef]
- Park, I.; Yang, S.; Kim, W.J.; Noh, P.; Lee, H.O.; Moon, B.C. The complete chloroplast genome of Cuscuta pentagona Engelm. Mitochondrial DNA B Resour. 2018, 3, 523–524. [Google Scholar] [CrossRef]
- Korea Institute of Oriental Medicine (KIOM). Defining Dictionary for Medicinal Herbs. 2019. Available online: http://boncho.kiom.re.kr/codex/ (accessed on 1 March 2019).
- Kim, E.-Y.; Kim, E.K.; Lee, H.-S.; Sohn, Y.; Soh, Y.; Jung, H.-S.; Shon, N.-W. Protective effects of Cuscutae semen against dimethylnitrosamine-induced acute liver injury in Sprague-Dawley rats. Biol. Pharm. Bull. 2007, 30, 1427–1431. [Google Scholar] [CrossRef]
- Kang, S.Y.; Jung, H.W.; Lee, M.-Y.; Lee, H.W.; Chae, S.W.; Park, Y.-K. Effect of the semen extract of Cuscuta chinensis on inflammatory responses in LPS-stimulated BV-2 microglia. Chin. J. Nat. Med. 2014, 12, 573–581. [Google Scholar] [PubMed]
- Liao, J.-C.; Chang, W.-T.; Lee, M.-S.; Chiu, Y.-J.; Chao, W.-K.; Lin, Y.-C.; Lin, M.-K.; Peng, W.-H. Antinociceptive and anti-inflammatory activities of Cuscuta chinensis seeds in mice. Am. J. Chin. Med. 2014, 42, 223–242. [Google Scholar] [CrossRef]
- Choi, B.H.; Cuscuta, L. The Genera of Vascular Plants of Korea; Flora of Korean Editorial Committee, Ed.; Academic Publishing Co.: Seoul, Korea, 2007; pp. 793–794. [Google Scholar]
- Park, S.H. New Illustrations and Photographs of Naturalized Plants of Korea; Ilchokak: Seoul, Korea, 2009. [Google Scholar]
- Kuijt, J. The Biology of Parasitic Flowering Plants; University of California Press: Berkeley, CA, USA, 1969. [Google Scholar]
- Choisy, J.D. De Convolvulaceis dissertatio tertia, complectens Cuscutarum hucusque cog nitarum methodicam enumerationem et descriptionem, necnon et brevem gallicam de Cuscutis praefationem. Mem. Soc. Phys. Hist. Nat. Genéve 1841, 9, 261–288. [Google Scholar]
- Engelmann, G. Systematic arrangement of the species of the genus Cuscuta with critical remarks on old species and descriptions of new ones. Trans. Acad. Sci. Saint Louis 1859, 1, 453–523. [Google Scholar]
- Peter, A. Convolvulaceae. In Die Natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; W. Engelmann: Leipzig, Germany, 1897. [Google Scholar]
- Mirande, M. Recherches Physiologiques et Anatomiques sur les Cuscutacées. Ph.D. Thesis, Faculté des Sciences de Paris, Paris, France, 1900. [Google Scholar]
- Hunziker, A.T. Las especies de Cuscuta (Convolvulaceae) de Argentina y Uruguay. Revista de la Facultad de Ciencias Exactas Fisicas y Naturales; Impr. de la Universidad: Córdoba, Argentina, 1950; pp. 1101–1202. [Google Scholar]
- Yuncker, T.G. Cuscuta. In North American Flora II. Part 4; New York Botanical Garden, Ed.; The New York Botanical Garden Press: New York, NY, USA, 1965; pp. 1–51. [Google Scholar]
- Wright, M.A.; Ianni, M.D.; Costea, M. Diversity and evolution of pollen-ovule production in Cuscuta (dodders, Convolvulaceae) in relation to floral morphology. Plant Syst. Evol. 2012, 298, 369–389. [Google Scholar] [CrossRef]
- García, M.A.; Costea, M.; Kuzmina, M.; Stefanović, S. Phylogeny, character evolution, and biogeography of Cuscuta (dodders; Convolvulaceae) inferred from coding plastid and nuclear sequences. Am. J. Bot. 2014, 101, 670–690. [Google Scholar] [CrossRef] [PubMed]
- Abdel Khalik, K.N. Seed morphology of Cuscuta L. (Convolvulaceae) in Egypt and its systematic significance. Feddes Repert. 2006, 117, 217–224. [Google Scholar] [CrossRef]
- Wang, Y.; Zhan, D.F.; Jia, X.; Mei, W.L.; Dai, H.F.; Chen, X.T.; Peng, S.Q. Complete chloroplast genome sequence of Aquilaria sinensis (Lour.) Gilg and evolution analysis within the Malvales order. Front. Plant Sci. 2016, 7, 280. [Google Scholar] [CrossRef]
- Wolfe, K.H.; Morden, C.W.; Palmer, J.D. Function and evolution of a minimal plastid genome from a nonphotosynthetic parasitic plant. Proc. Natl. Acad. Sci. USA. 1992, 89, 10648–10652. [Google Scholar] [CrossRef]
- Kim, H.T.; Kim, J.S.; Moore, M.J.; Neubig, K.M.; Williams, N.H.; Whitten, W.M.; Kim, J.H. Seven new complete plastome sequences reveal rampant independent loss of the ndh gene family across Orchids and associated instability of the inverted repeat/small single-copy region boundaries. PLoS ONE 2015, 10, e0142215. [Google Scholar] [CrossRef]
- Wicke, S.; Muller, K.F.; dePamphilis, C.W.; Quandt, D.; Bellot, S.; Schneeweiss, G.M. Mechanistic model of evolutionary rate variation en route to a nonphotosynthetic lifestyle in plants. Proc. Natl. Acad. Sci. USA 2016, 113, 9045–9050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Z.; Ye, Y.; Bai, T.; Xu, M.; Xu, L.A. Complete chloroplast genome of Pinus massoniana (Pinaceae): Gene rearrangements, loss of ndh genes, and short inverted repeats contraction, expansion. Molecules 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; McDevitt, R.; Vendramin, G.G.; Rafalski, J.A. Polymorphic simple sequence repeat regions in chloroplast genomes: Applications to the population genetics of pines. Proc. Natl. Acad. Sci. USA 1995, 92, 7759–7763. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Cheng, T.; Lin, K.; Zhou, S. Sequencing angiosperm plastid genomes made easy: A complete set of universal primers and a case study on the phylogeny of Saxifragales. Genome Biol. Evol. 2013, 5, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Lv, S.; Zhang, Y.; Du, X.; Wang, L.; Biradar, S.S.; Tan, X.; Wan, F.; Weining, S. Complete chloroplast genome sequence of a major invasive species, crofton weed (Ageratina adenophora). PLoS ONE 2012, 7, e36869. [Google Scholar] [CrossRef] [PubMed]
- Khakhlova, O.; Bock, R. Elimination of deleterious mutations in plastid genomes by gene conversion. Plant J. 2006, 46, 85–94. [Google Scholar] [CrossRef]
- Hansen, D.R.; Dastidar, S.G.; Cai, Z.; Penaflor, C.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Phylogenetic and evolutionary implications of complete chloroplast genome sequences of four early-diverging angiosperms: Buxus (Buxaceae), Chloranthus (Chloranthaceae), Dioscorea (Dioscoreaceae), and Illicium (Schisandraceae). Mol. Phylogenet. Evol. 2007, 45, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Shi, C.; Liu, Y.; Mao, S.Y.; Gao, L.Z. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: Genome structure and phylogenetic relationships. BMC Evol. Biol. 2014, 14, 151. [Google Scholar] [CrossRef] [PubMed]
- Braukmann, T.; Kuzmina, M.; Stefanovic, S. Plastid genome evolution across the genus Cuscuta (Convolvulaceae): Two clades within subgenus Grammica exhibit extensive gene loss. J. Exp. Bot. 2013, 64, 977–989. [Google Scholar] [CrossRef]
- Moore, M.J.; Bell, C.D.; Soltis, P.S.; Soltis, D.E. Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms. Proc. Natl. Acad. Sci. USA 2007, 104, 19363–19368. [Google Scholar] [CrossRef] [Green Version]
- The Angiosperm Phylogeny Group; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Bell, C.D.; Edwards, E.J.; Kim, S.-T.; Donoghue, M.J. Dipsacales phylogeny based on chloroplast DNA sequences. Harv. Pap. Bot. 2001, 6, 481–499. [Google Scholar]
- Song, J.-H.; Moon, B.C.; Choi, G.; Yang, S. Morphological identification of Lepidii Seu Descurainiae Semen and adulterant seeds using microscopic analysis. Appl. Sci. 2018, 8, 2134. [Google Scholar] [CrossRef]
- Delcher, A.L.; Salzberg, S.L.; Phillippy, A.M. Using MUMmer to identify similar regions in large sequence sets. Curr. Protoc. Bioinform. 2003, 1, 10.3.1–10.3.18. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T. MISA—Microsatellite identification tool. 2003. Available online: Website http://misaweb.ipk-gatersleben. de/. (accessed on 2 March 2019).
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef]
- Warburton, P.E.; Giordano, J.; Cheung, F.; Gelfand, Y.; Benson, G. Inverted repeat structure of the human genome: The X-chromosome contains a preponderance of large, highly homologous inverted repeats that contain testes genes. Genome Res. 2004, 14, 1861–1869. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
Sample Availability: Samples of Cuscuta japonica and C. chinensis are available from the authors and the herbarium of KIOM. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. japonica | C. reflexa | C. exaltata | C. chinensis | C. gronovii | C. pentagona | C. obtusiflora | |
|---|---|---|---|---|---|---|---|
| GenBank accession number | This study | NC009766 | NC009963 | This study | NC009765 | NC039759 | NC009949 |
| Chloroplast genome size (bp) | 121,037 | 121,521 | 125,373 | 86,927 | 86,744 | 86,380 | 85,280 |
| Large single copy (LSC) region (bp) | 79,517 | 79,468 | 82,721 | 50,572 | 50,973 | 50,958 | 50,201 |
| Inverted repeat (IR) region (bp) | 16,554 | 16,741 | 16,701 | 14,617 | 14,354 | 14,200 | 14,131 |
| Small single copy (SSC) region (bp) | 8412 | 8571 | 9250 | 7121 | 7063 | 7022 | 6817 |
| Total number of genes | 96 | 98 | 98 | 85 | 86 | 85 | 86 |
| Number of protein-coding genes | 65 | 65 | 65 | 58 | 58 | 57 | 58 |
| Number of rRNA genes | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| Number of tRNA genes | 27 | 29 | 29 | 23 | 24 | 24 | 24 |
| GC content (%) | 38.3 | 38.2 | 38.1 | 37.6 | 37.7 | 37.9 | 37.8 |
| LSC (%) | 36 | 36.1 | 35.8 | 35.9 | 35.8 | 36 | 35.9 |
| IR (%) | 45.7 | 45.7 | 45.6 | 42.8 | 43.1 | 43.2 | 43.2 |
| SSC (%) | 30.5 | 30.8 | 31 | 28.9 | 29.5 | 29.6 | 29.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, I.; Song, J.-H.; Yang, S.; Kim, W.J.; Choi, G.; Moon, B.C. Cuscuta Species Identification Based on the Morphology of Reproductive Organs and Complete Chloroplast Genome Sequences. Int. J. Mol. Sci. 2019, 20, 2726. https://doi.org/10.3390/ijms20112726
Park I, Song J-H, Yang S, Kim WJ, Choi G, Moon BC. Cuscuta Species Identification Based on the Morphology of Reproductive Organs and Complete Chloroplast Genome Sequences. International Journal of Molecular Sciences. 2019; 20(11):2726. https://doi.org/10.3390/ijms20112726
Chicago/Turabian StylePark, Inkyu, Jun-Ho Song, Sungyu Yang, Wook Jin Kim, Goya Choi, and Byeong Cheol Moon. 2019. "Cuscuta Species Identification Based on the Morphology of Reproductive Organs and Complete Chloroplast Genome Sequences" International Journal of Molecular Sciences 20, no. 11: 2726. https://doi.org/10.3390/ijms20112726