
Tolerance and accumulation of lithium in Apocynum pictum Schrenk
- Published
- Accepted
- Received
- Academic Editor
- Todd Anderson
- Subject Areas
- Ecology, Plant Science, Environmental Contamination and Remediation
- Keywords
- Bioconcentration factor, Apocynum pictum, Lithium, Phytoremediation, Germination, Translocation factor
- Copyright
- © 2018 Jiang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. Tolerance and accumulation of lithium in Apocynum pictum Schrenk. PeerJ 6:e5559 https://doi.org/10.7717/peerj.5559
Abstract
Primarily, lithium (Li) resource development and wider application of Li-ion batteries result in Li pollution and concomitantly poses increasing and inevitable problems to environmental health and safety. However, information is rare about the scope of the remediation of Li contaminated soil. Apocynum venetum is already proved to be a Li-accumulator with high Li tolerance and accumulation (Jiang et al., 2014). However, it is not clear whether Apocynum pictum, another species of the same genus with the same uses as A. venetum, is also a Li-accumulator. We investigated germination, growth and physiological responses of A. pictum to different levels of LiCl. Germination was not significantly affected by low Li concentration (0–100 mmol L−1). As LiCl increased from 100 to 400 mmol L−1, both germination percentage and index decreased gradually. For germination of A. pictum seeds, the critical value (when germination percentage is 50%) in LiCl solution was 235 mmol L−1, and the limit value (when germination percentage is 0%) was 406 mmol L−1. A. pictum could accumulate >1,800 mg kg−1 Li in leaves, and still survived under 400 mg kg-1 Li supply. The high Li tolerance of A. pictum during germination and growth stage was also reflected by activity of α-amylase and contents of soluble sugar, proline and photosynthetic pigments under different Li treatments. The bioconcentration factors (BCF) (except control) and translocation factors (TF) were higher than 1.0. High tolerance and accumulation of Li indicated that A. pictum is Li-accumulator. Therefore, this species could be useful for revegetation and phytoremediation of Li contaminated soil.
Introduction
Lithium (Li), the lightest alkali metal, is normally present in trace amounts in the environment (Shahzad et al., 2017). As the beneficial and detrimental effects to living organisms depending on the dose of Li, more attention should be paid to the increase of Li contents in soil caused by industrial activities, use and disposal of Li-containing products (Schrauzer, 2002). The increase of toxic hazard is mainly caused by the dispose of used-up Li-ion batteries (Aral & Vecchio-Sadus, 2008; Robinson et al., 2018). Li is listed as a pollutant that causes environmental harm in irrigation water supplies (Australian Capital Territory Parliamentary Counsel, 2005).
In the consideration of Li toxicity and its impact on health and environment, several remediation methods have been proposed to reduce its toxicity and remediate Li contaminated soil. These approaches include: (1) identification and use of Li tolerant genotypes, accumulator and hyperaccumulator; (2) organic matter addition to soils; (3) application of chelators or chaperones; (4) Ex-situ and in-situ immobilization techniques; (5) soil stabilization and solidification (Shahzad et al., 2016). Among these methods, phytoremediation is inexpensive and can reduce secondary pollution compared with traditional physical and chemical approaches (Cluis, 2004; Franzaring et al., 2016). However, lack of necessary information about Li accumulator and hyperaccumulator is the limitation factor of this technology. Screening some Li-accumulating plant species is the first step to remediate Li-contaminated soil using phytoremediation.
Although Li is a non-essential metal for plant growth, the growth-stimulating effects of Li at low concentration are reported. For example, fresh biomass was increased by 15% in maize treated with 5 mg L−1 Li (Hawrylak-Nowak, Kalinowska & Szymańska, 2012). Meanwhile, Li induces considerable reduction in the growth of plants as the Li-salts are highly toxic (Franzaring et al., 2016). Citrus plants are sensitive to Li (Aral & Vecchio-Sadus, 2008). However, plants in the Asteraceae and Solanaceae families have high tolerance to Li (Schrauzer, 2002; Kabata-Pendias & Mukherjee, 2007). Thus, Li influences plant growth in both stimulation and reduction ways, depending on Li concentration and plant species.
Apocynum pictum, perennial halophytic herb, mainly distributes in salt-barren zone, desert margins, and riversides of the northwestern region of China (Flora of China Editorial Committee, 1995). Like Apocynum venetum, another species of the same genus, A. pictum is also a popular daily beverage and commonly used as traditional Chinese medicine to calm the liver, sooth nerves, and promote dieresis (Thevs et al., 2012). Modern pharmacological studies have demonstrated that many pharmacological effects of both plants are similar to those of Li (Editorial Committee of Chinese Pharmacopoeia, 2010; Khasraw et al., 2012). A. venetum is already proved to be a Li-accumulator (Jiang et al., 2014), so we hypothesized that A. pictum is also a Li-accumulator.
The aim of this study was to examine the accumulation capacity of Li in A. pictum and also to examine Li toxicity tolerance ability. Specifically, three questions were addressed: (1) What is Li tolerance for A. pictum at germination stage? (2) Does an increase in soil Li contents inhibit the growth of A. pictum and reduce photosynthetic pigments? (3) Are Li contents in different organs of A. pictum affected by elevated soil Li?
Materials and Methods
Germination experiment
Mature fruits were collected from plants of A. pictum cultivated at Fukang Field Research Station of the Chinese Academy of Sciences (44°17′N, 87°56′E; 460 m a.s.l.), located in the southern part of the Junggar Basin of Xinjiang Province, China, in early October 2016. Fruits were allowed to dry naturally for two weeks. Then, the seeds were detached from fruits and stored dry at room temperature.
Germination experiment was performed on 4th November 2016. Seeds were placed on two layers of filter paper in 5-cm-diameter Petri dishes. Filter paper was moistened with 2.5 mL of 0 (distilled water), 25, 50, 100, 150, 200, 300 and 400 mmol L−1 LiCl (Tianjin Fuchen Chemical Reagent Co., Ltd). Four replicates of 25 seeds were used per treatment. The lids were sealed with Parafilm and incubated at 25 °C under a 14/10 h light/dark photoperiod for 15 days. Lithium solutions were changed every 5th day. The criteria for germination were the radicle length ≥1 mm. The number of germinated seeds was counted and germinated seeds were removed from the Petri dishes at each counting. The velocity of germination was estimated using a modified Timson’s index of germination velocity (Khan & Ungar, 1984).
Physiological parameter
Seeds of A. pictum were incubated on two layers of filter paper in 5-cm-diameter Petri dishes. Filter paper was moistened with 0 (distilled water control), 25, and 300 mmol L−1 LiCl. Petri dishes were sealed with parafilm and incubated at 25 °C under a 14/10 h light/dark photoperiod. Incubation was terminated after 2, 4, 6, 8, 10 or 12 d.
To extract α-amylase, fresh seeds (0.2 g) were homogenized with 8.0 mL distilled water. Activity of α-amylase was measured spectrophotometrically at 540 nm (Li, 2000). After fresh seeds (0.2 g) were homogenized with 10 mL of 10% trichloroacetic acid, the malondialdehyde (MDA) content was determined according to the method of (Duan et al., 2005). After fresh seeds (0.2 g) were homogenized with 10.0 mL distilled water, soluble sugar content was measured spectrophotometrically at 630 nm (Li, 2000). After fresh seeds (0.2 g) were homogenized with 5.0 mL of 3% sulfonyl salicylic acid, proline content was measured spectrophotometrically at 520 nm (Li, 2000).
Pot experiment
The pots were placed in a greenhouse at Fukang Field Research Station of the Chinese Academy of Sciences. Gray desert topsoil (0–20 cm) was collected and used as the growth substrate. The soil was air-dried and homogenized, after which it was sieved through 4-mm mesh. Soil pH was 8.47, organic matter 0.92 %, total N 0.96 g kg−1, available N 89.54 mg kg−1, available P 9.24 mg kg−1, available K 341.16 mg kg−1, and Li 25.75 mg kg−1. Each plastic pot (35 cm height, 33 cm diameter) was filled with 10 kg of air-dry soil mixed with different contents of Li (solution prepared by dissolving analytical grade LiCl): 0 (CK without external Li), 50, 200, and 400 mg kg−1. To simulate the Li impacted soils, pot soil was homogenized for four weeks with the additive Li. Two A. pictum rhizome cuttings of similar size (9–10 cm height, ca. 1 cm diameter) were transplanted into each pot on 14 April, 2012. There were eight replicates for each treatment, and they were arranged in a randomized block design. Each pot received 10 g granular lawn fertilizer (Osmocote 301; Scotts, Marysville, OH, USA; 15N: 11P: 13K: 2Mg) once at the beginning as the basic fertilizer. Each pot was irrigated with 1 L distilled water every two day.
After 90 days of cultivation, A. pictum plants were harvested and separated into roots, stems, and leaves. The leaves and stems were rinsed with distilled water three times and then blotted with filter paper. The roots were dipped in a CaCl2 solution for 5 minutes to get rid of trace elements adsorbed at the root surface and then rinsed three times with distilled water. Fresh material was oven-dried at 80 °C for 48 h. Afterwards the samples were weighed, ground, and sieved through 60-mesh.
Photosynthetic pigments were extracted in darkness at 4 °C with 5 ml mixture of acetone and ethanol (v/v = 1:1) for 48 h. After filtering, chlorophyll a (Chl a), chlorophyll b (Chl b) and carotenoid (Car) contents were determined by spectrophotometry using 663, 645, and 470 wavelength. Pigment concentrations were calculated following Lichtenthaler (1987).
0.2 g plant sample was digested with 10 ml concentrated HNO3 (50%) for 12 h in triangular flask. Then, 3 ml H2O2 (30%) was added and kept in a temperature control electric cooker (Lab Tech EH35A) at 150 °C for 10 min. After cooled to 20 °C, 5 ml HNO3 (1%) and 1ml H2O2 (30%) were added. Sample was digested at 150 °C until the white smoke emitted fully and the solution was transparent. After cooled to 20 °C, the solution was diluted to 50 ml with HNO3 (1%). Li content was estimated with an inductively coupled plasma spectroscopy (Perkin Elmer SciexDRC II) in the range of 670–783 nm (Kalinowska, Hawrylaknowak & Szymańska, 2013), and determined at 670.783 nm.
Bioconcentration factors (BCF) is defined as the ratio of the metal concentration in the root to that in the soil; Translocation factors (TF) is defined as the ratio of the metal concentration in the shoot to that in the root (Baker & Brooks, 1989).
Statistical analysis
Germination percentage data were arcsine transformed before statistical analysis. One-way ANOVA was used to test the effect of different Li concentrations on germination, dry weight, pigment contents, Li concentrations, BCF, and TF. Duncan’s test was used to determine differences among treatments when ANOVA showed significant effects (P < 0.05).
Results
Germination characteristics of A. pictum seeds
Germination percentage and index of A. pictum seeds were significantly affected by LiCl. With the increase of concentration of LiCl, germination percentages and indexes decreased (Fig. 1). Germination percentages of A. pictum seeds were reduced from 91% to 6% under 0–400 mmol L−1 LiCl (Fig. 1A). Under low to moderate concentration of LiCl (25–100 mmol L−1), A. pictum seeds achieved high germination percentage (>80%). Even under 200 mmol L−1 LiCl, germination percentage could reach 54%. The regression equation of the relationship between germination percentages of A. pictum seeds and LiCl concentrations is: y = − 0.0002777x2 − 0.1141x + 92.03 (R2 = 0.9538). The simulated critical value (when germination percentage is 50%) is 235 mmol L−1. The simulated limit value (when germination percentage is 0%) is 406 mmol L−1.
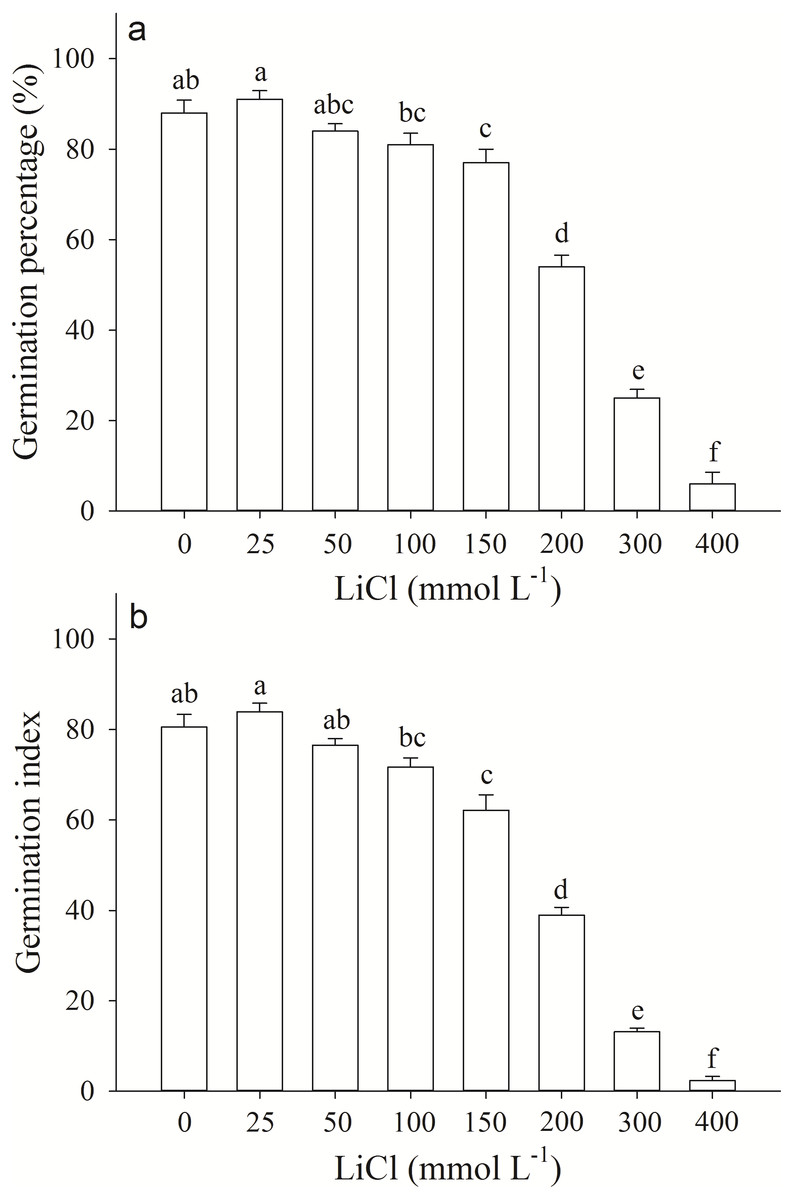
Figure 1: Germination response of Apocynum pictum seeds to LiCl.
Effects of LiCl solution on germination percentage (A) and germination index (B) of Apocynum pictum seeds. Different letters indicate significant differences in germination percentages or germination index among different LiCl concentrations with Duncan’s test (P < 0.05).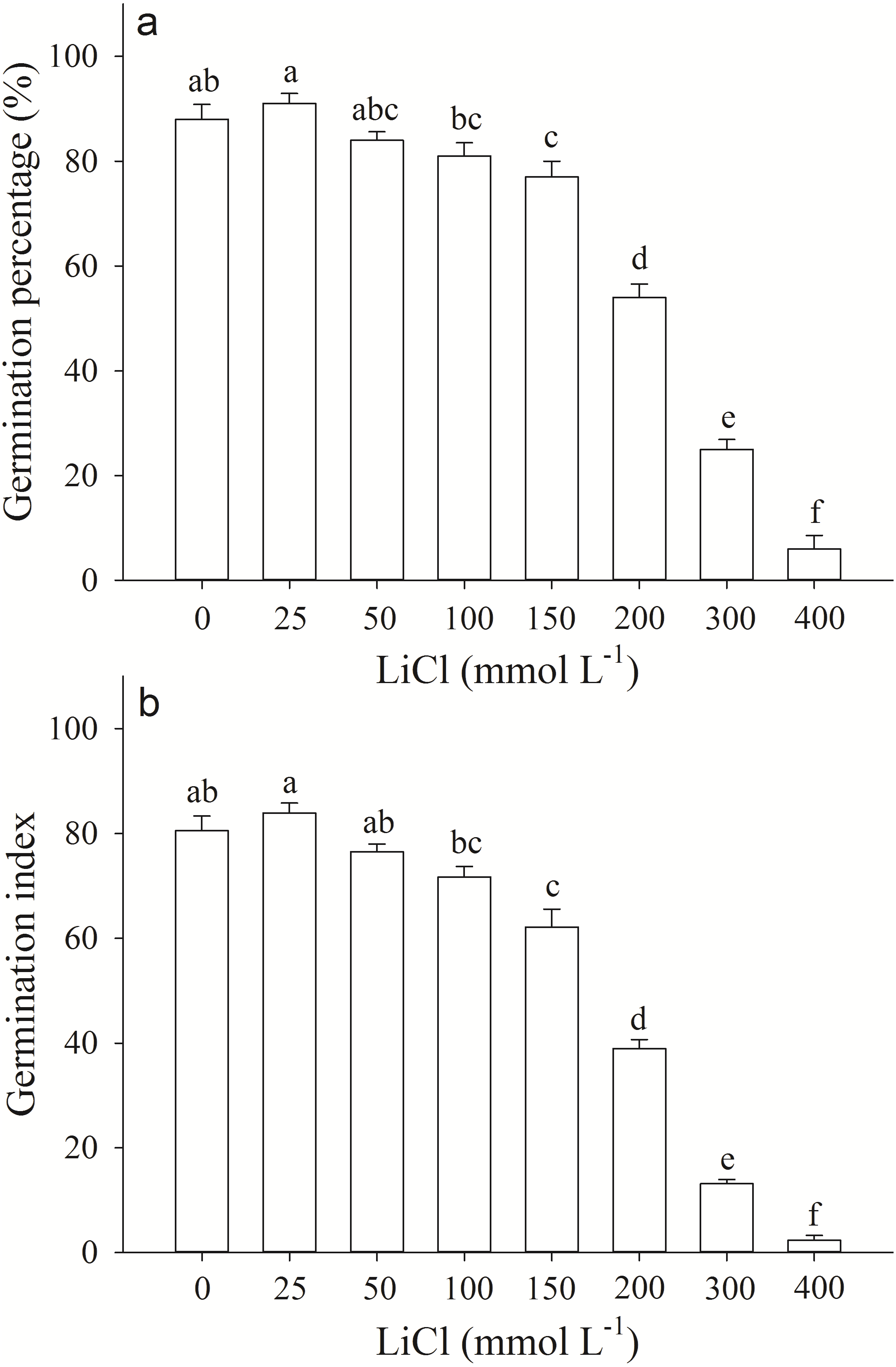{kind=link}
Activity of α-amylase, contents of MDA, soluble sugar and proline were dramatically affected by LiCl (Fig. 2). Least α-amylase activity was noted under 300 mmol L−1 LiCl (Fig. 2A). Contents of MDA, soluble sugar and proline under 300 mmol L−1 LiCl were the highest (Figs. 2B–2D). Activity of α-amylase fist increased and then decreased under Li treatment and was highest after 6 days of incubation. Contents of MDA increased with treated time in LiCl.

Figure 2: Physiological parameters of Apocynum pictum to LiCl during germination.
Effects of LiCl concentration on (A) α-amylase activity, (B) MDA, (C) soluble sugar and (D) proline contents in seeds of Apocynum pictum during germination.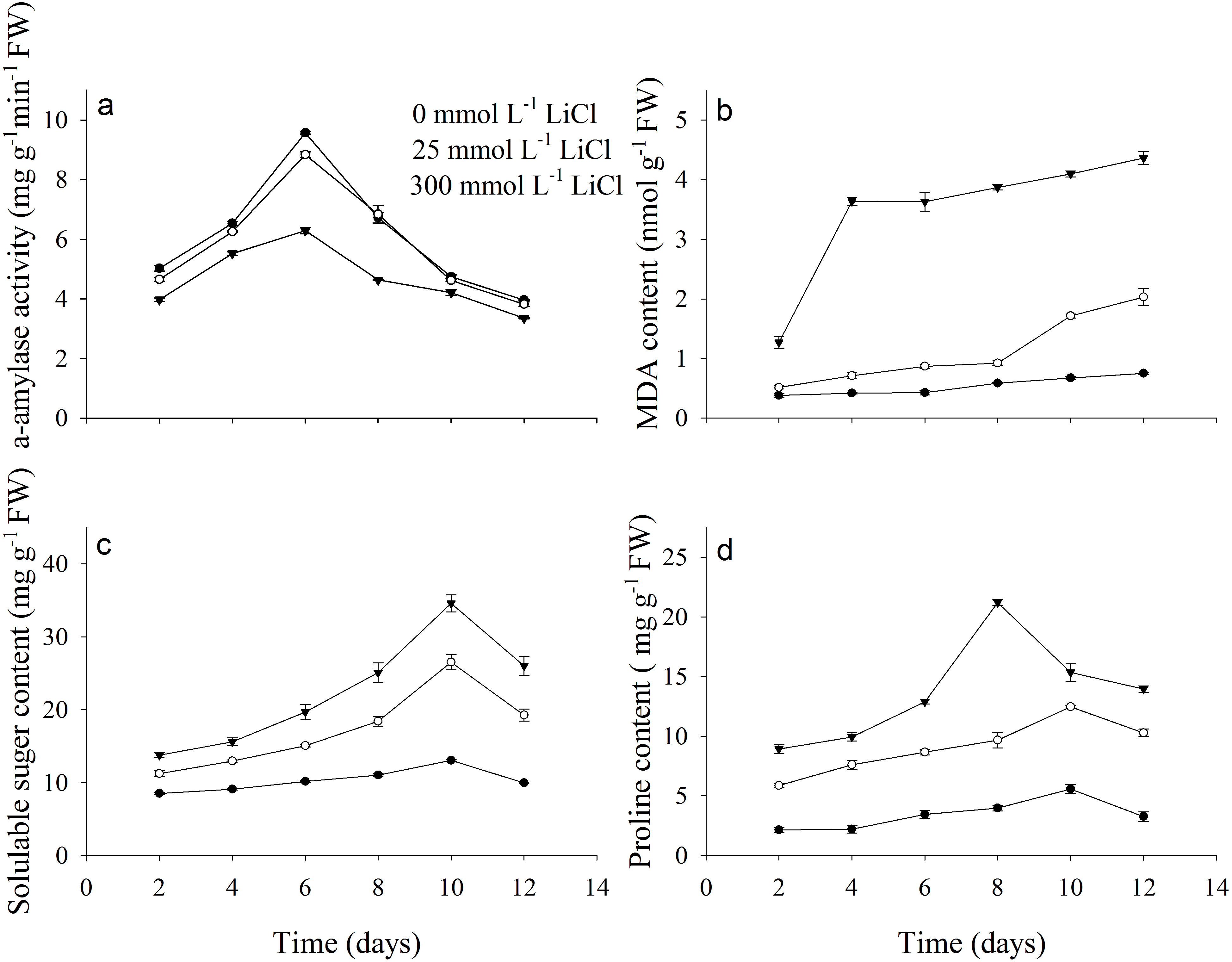{kind=link}
Growth and pigment contents of A. pictum plants
Dry weight of leaf, stem and root of A. pictum did not significantly change at 50 mg kg−1 additional soil Li, but only significantly decreased at 200 and 400 mg kg−1 additional Li (Figs. 3A, 3C, 3E). When compared with 0 mg kg−1 Li, the reductions of leaves dry weight were 42.90% and 78.15% under 200 and 400 mg kg−1 Li treatment, respectively; the reductions of stems dry weight were 45.34% and 74.02% under 200 and 400 mg kg−1 Li compared with control respectively; the reductions of roots dry weight were 40.41% and 64.02% under 200 and 400 mg kg−1 Li treatment respectively.

Figure 3: Growth and physiological responses of Apocynum pictum to additional soil Li.
Effects of additional soil Li on leaf dry weight (A), stem dry weight (C), root dry weight (E), chlorophyll a content (B), chlorophyll b content (D), and car content (F) of Apocynum pictum. Different letters indicate significant differences in each measures among different additional soil Li contents with Duncan’s test (P < 0.05).{kind=link}
Pigment contents (Chl a, Chl b, and Car) were not significantly affected by 50 mg kg−1 additional Li (Figs. 3B, 3D & 3F). However, for plants grown at 200 and 400 mg kg−1 additional Li, there were significant reductions in contents of Chl a, Chl b, and C ar compared with control. When compared with 0 mg kg−1 Li, the reductions of Chl a contents were 29.96% and 78.30% under 200 and 400 mg kg−1 Li treatment, respectively; the reductions of Chl b contents were 24.09% and 76.75% under 200 and 400 mg kg−1 Li respectively; the reductions of Car contents were 32.49% and 76.75% under 200 and 400 mg kg−1 Li respectively.
Li accumulation and translocation of A. pictum plants
Li accumulation in different plant tissues were significantly influenced by external Li concentration in soil, however among different tissues, Li concentration in leaves was significantly higher than that in stems and roots (P < 0.05; Fig. 4). Moreover, in leaves and stem tissues, Li concentration was increased, marked from 0 to 200 mg kg−1 additional Li (P < 0.05), and then deceased from 200 to 400 mg kg−1 additional Li. Contrarily, in A. pictum roots, Li concentration was proportionally increased with an increase in additional Li in soil (0 to 400 mg kg−1) (Fig. 4). When compared with Li contents in leaves, the decrements of Li contents in roots were 39.62%, 86.80%, 75.60%, and 61.82% under 0, 50, 200 and 400 mg kg−1 Li treatment, respectively.
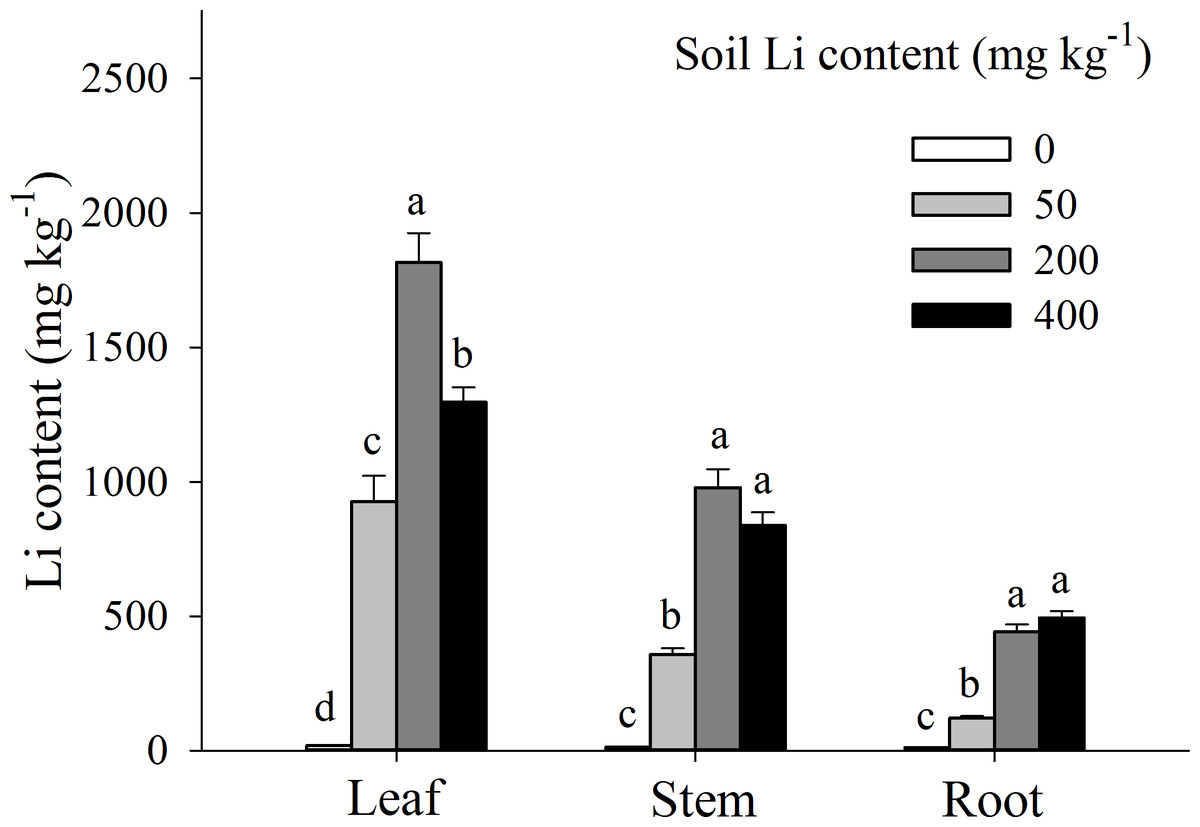
Figure 4: Li contents in different organs of Apocynum pictum in responses to additional soil Li.
Effects of additional soil Li on Li contents in leaf, stem and root of Apocynum pictum. Different letters indicate significant differences in Li contents of the same plant organ with Duncan’s test (P < 0.05).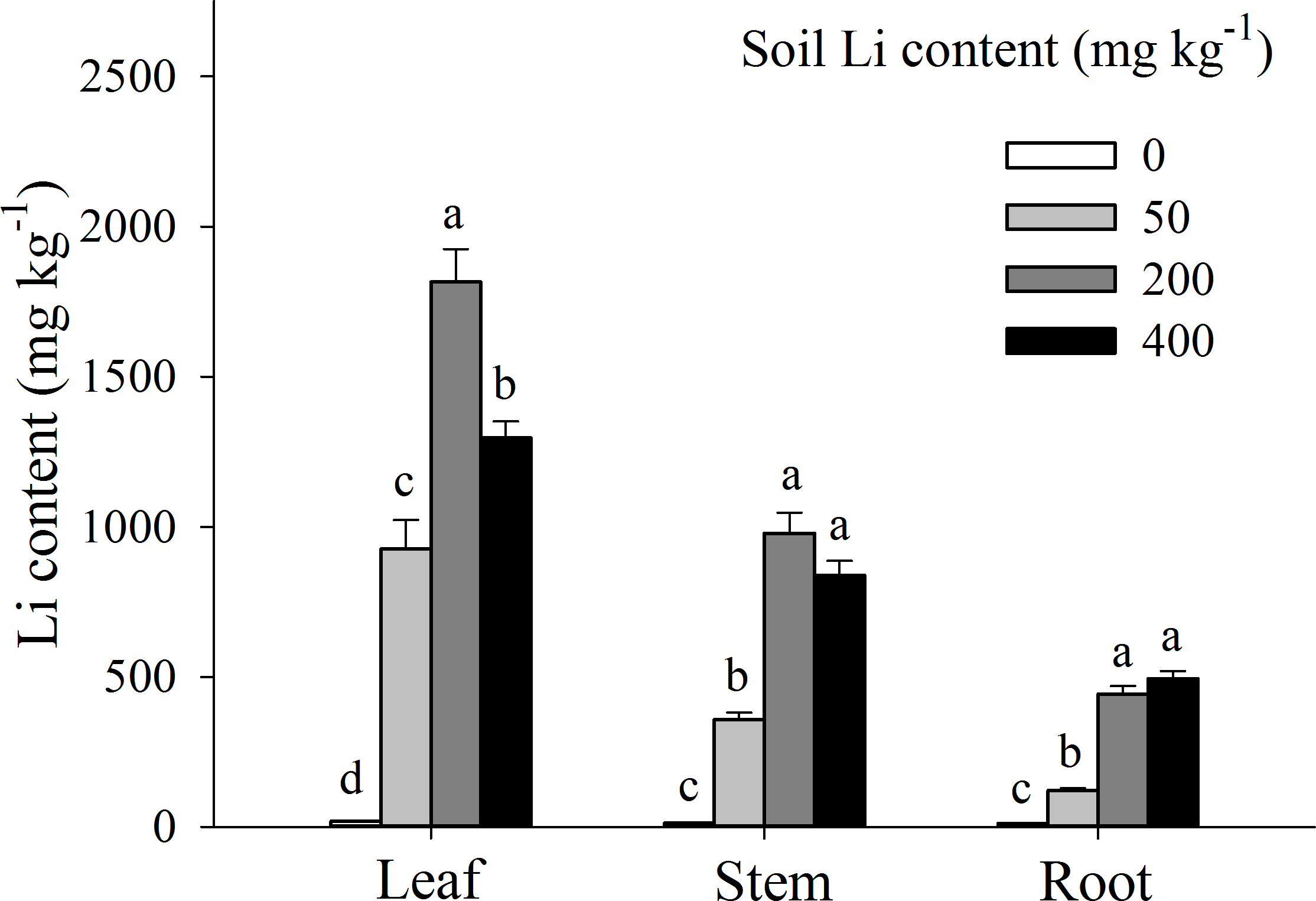{kind=link}
The bioconcentration factors (BCF) and translocation factors (TF) values were greater than 1.0 for most Li treatments (except control). After 90 d of cultivation, BCF value of A. pictum at 0 (control), 50, 200, and 400 mg kg−1 additional Li was 0.45, 1.61, 1.96, 1.16, respectively (Fig. 5A). TF value at 0, 50, 200, and 400 mg kg−1 additional Li was 1.42, 5.24, 3.30, 2.15, respectively (Fig. 5B).
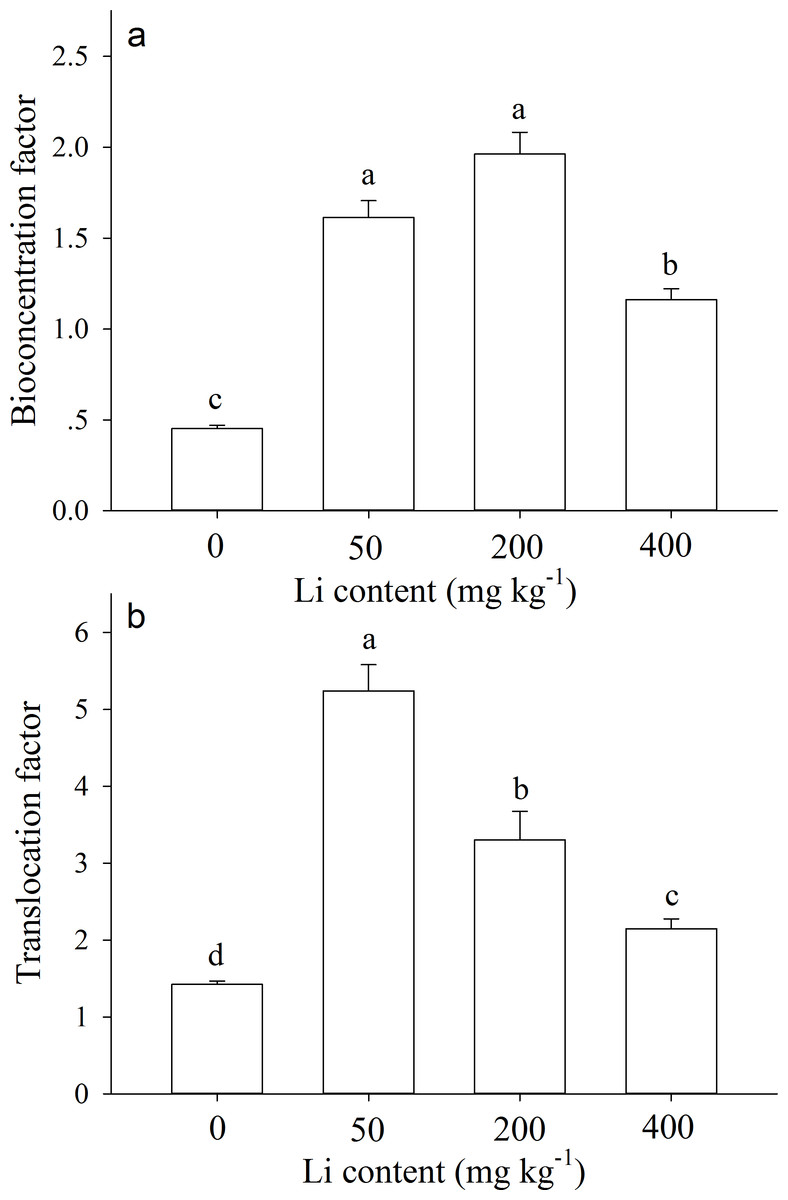
Figure 5: Bioconcentration factor and translocation factor of Apocynum pictum to additional soil Li.
Effects of additional soil Li on bioconcentration factor (BCF) (A) and translocation factor (TF) (B) of Apocynum pictum. Different letters indicate significant differences in BCF or TF among different additional soil Li contents with Duncan’s test (P < 0.05).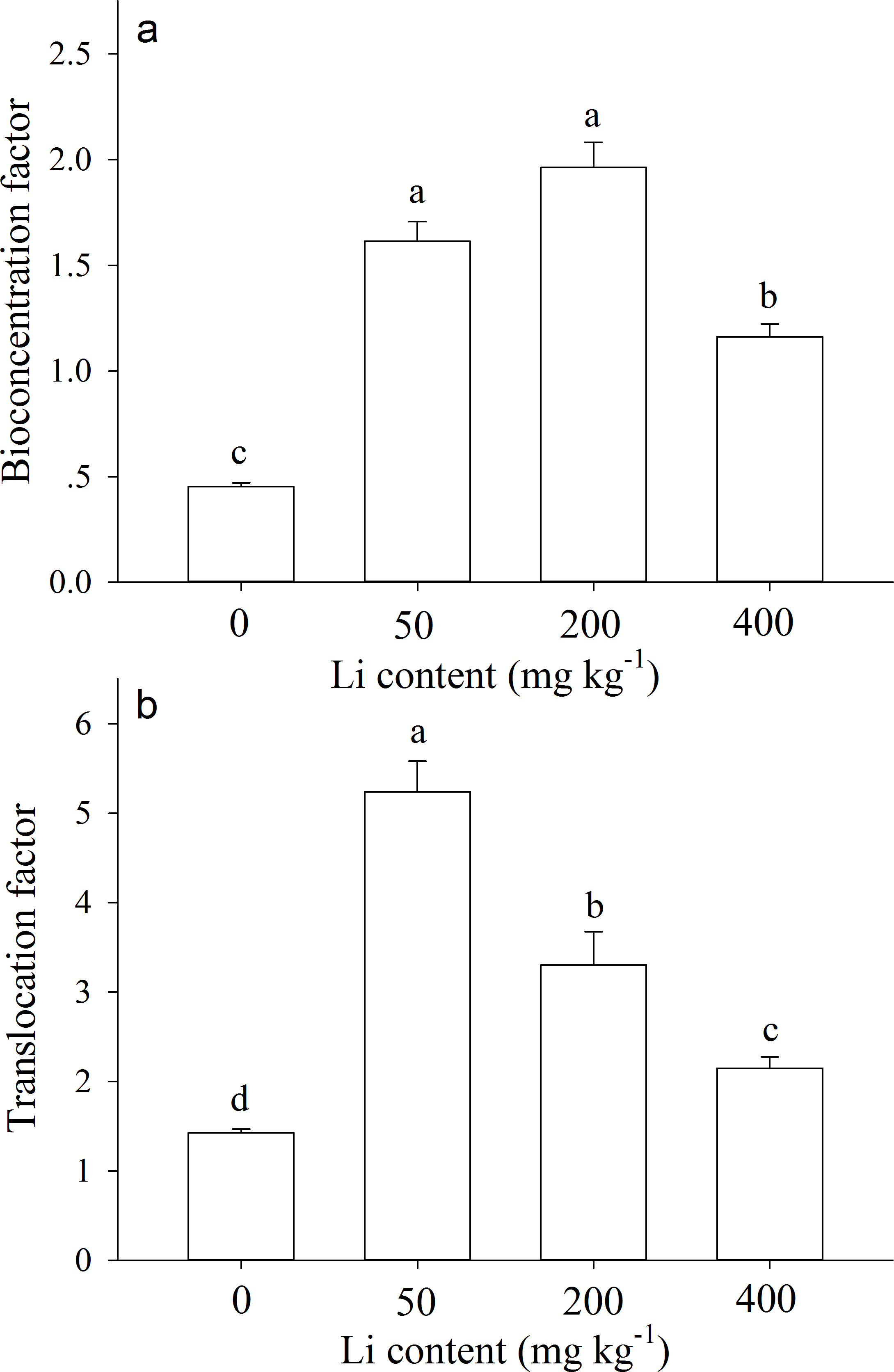{kind=link}
Discussion
Although germination, plant growth, and ecophysiological responses of A. pictum to environmental factors have been studied extensively (Ma et al., 2003; Thevs et al., 2012; Shi et al., 2014), these data are the first in which tolerance and accumulation characteristics of Li in A. pictum plants have been documented. In addition, our data indicate that seeds of A. pictum have high Li tolerance during germination. Our results of plant growth and Li accumulation traits indicated A. pictum as a potential Li-accumulator.
Cultivation of Li-accumulator in Li polluted soil might be an economical and eco-friendly method for revegetation (Shahzad et al., 2016). The information of germination responses of Li-accumulator seeds to different Li concentrations is vital for soil Li phytoremediation purpose. For Li-sensitive plants, such as Oryza sativa and Cicer arietinum, germination percentages were affected by 0.72 mmol L−1 (Gupta, 1974). Even for potential Li accumulator Brassica carinata, germination percentage was <50% at 120 mmol L−1 LiCl (Li et al., 2009). For Li-accumulator Apocynum venetum, the critical value (when germination percentage is 50%) in LiCl solution is 196 mmol L−1 (Jiang, Wang & Tian, 2018). Our results showed germination percentage of A. pictum was 54% at 200 mmol L−1 LiCl, and showed that A. pictum exhibited high Li tolerance during germination stage (Fig. 1).
The high Li tolerance of A. pictum during germination stage was also reflected by different physiological parameters. α-amylase plays an important role in the starch metabolism during germination (Muralikrishna & Nirmala, 2005). Activity of α-amylase is an indicator of catalytic efficiency of hydrolysis of starch into sugars (Salieri, Vinci & Antonelli, 1995). There was no significantly difference in the activity of α-amylase under 0 and 25 mmol L−1 LiCl. This result indicated the 25 mmol L−1 LiCl did not dramatically affect the changes in α-amylase involved in starch breakdown during germination compared with 0 mmol L−1 Li treatment. Organic solutes play an important role in osmotic adjustment in plants under salt stress (Martino et al., 2003). A. pictum seeds during germination under different Li treatments accumulated high concentrations of soluble sugar and proline (Fig. 2). This clearly highlighted that A. pictum can tolerate high Li concentration by increasing the accumulation contents of organic solutes. A. pictum accumulated proline and soluble sugar as cheap osmoticum to ameliorate Li induced toxicity. Further studies are required to investigate molecular basis of Li tolerance mechanisms in A. pictum.
Plants of A. pictum showed high tolerance to Li stress because plant growth and pigment contents were not significantly affected by 75 mg kg−1 soil Li. For Li tolerant species Chloris gayana and Zea mays, their growth were reduced up to 25% at 65 and 70 mg kg−1 soil Li, respectively (Bingham, Bradford & Page, 1964). Among different plant tissues, A. pictum roots accumulated small amount of Li, however, stems and leaves accumulated large amount of Li (Fig. 4). When additional soil Li reached 200 mg kg−1, Li content in stems and leaves of A. pictum was more than 950 and 1,800 mg kg−1, respectively (Fig. 4). Reported Li-accumulators, such as Cirsium arvense and Solanum dulcamera could accumulate 1,000 mg kg−1 Li in leaves (Tölgyesi, 1983). Compared with these plants, the capacity of A. pictum plants to accumulate Li is even better. Even compared with A. venetum, a newly found Li-accumulator of the same genus (Jiang et al., 2014), the capacity of A. pictum plants to tolerate and accumulate Li is equal. BCF and TF are key indexes used to evaluate metal accumulation efficiency in plants. BCF of A. pictum were 1.16–1.96 for 50–400 mg kg−1 additional soil Li, and TF of A. pictum were 2.15–5.24 for 50–400 mg kg−1 additional soil Li, respectively, which were higher than 1.0 (the critical level for accumulator).
Conclusions
Current evidence shows that A. pictum seeds have high Li tolerance and A. pictum plants have high Li tolerance and accumulation. Growth and physiological results confirm that A. pictum is a tolerant species to Li. Contents of Li in roots, stems and leaves, BCF and TF indicate that this species has the basic traits of a Li-accumulator. High Li tolerance of A. pictum seeds and plants implies that sowing this species directly is possible in Li contaminated soil and may be useful for phytoremediation and revegetation of Li contaminated sites. However, it is not clear whether other environmental factors such as soil physicochemical properties have a positive or negative effect in determining Li resistance and absorbing. Future research on multiple factor experiments will help to answer this question.
Supplemental Information
Germination data of Apocynum pictum to different LiCl concentrations
Four replicates of 25 seeds germinated under 0 (distilled water), 25, 50, 100, 150, 200, 300 and 400 mmol L −1 LiCl. The number of germinated seeds was counted every day. Each data point indicates the number of germinated seeds.
Physiological parameters of Apocynum pictum to different LiCl concentrations during germination
Four replicates of 25 seeds germinated under 0 (distilled water), 100 and 300 mmol L −1 LiCl. The physiological parameters of germinated seeds was tested every two days. Each data point indicates the physiological index of germinated seeds.
Biomass, contents of photosynthetic pigments and Li accumulation in different organs of Apocynum pictum
Each data point indicates weight, contents of photosynthetic pigments or Li in different organs of Apocynum pictum. Eight replicates for each treatment.