











Little Grebe Tachybaptus ruficollis Scientific name definitions
Text last updated May 10, 2014
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Kleindobbertjie |
| Albanian | Kredharaku i vogël |
| Arabic | غطاس صغير |
| Armenian | Փոքր սուզակ |
| Assamese | পানীডুবি |
| Asturian | Somorguyñn comñn |
| Azerbaijani | Kiçik batağan |
| Bangla | পানডুবি |
| Basque | Txilinporta txikia |
| Bulgarian | Малък гмурец |
| Catalan | cabusset comú |
| Chinese | 小鸊鷉 |
| Chinese (Hong Kong SAR China) | 小鸊鷉 |
| Chinese (SIM) | 小䴙䴘 |
| Croatian | mali gnjurac |
| Czech | potápka malá |
| Danish | Lille Lappedykker |
| Dutch | Dodaars |
| English | Little Grebe |
| English (Philippines) | Little Grebe |
| English (South Africa) | Dabchick (Little Grebe) |
| English (United States) | Little Grebe |
| Faroese | Smágjør |
| Finnish | pikku-uikku |
| French | Grèbe castagneux |
| French (France) | Grèbe castagneux |
| Galician | Merguillete común |
| German | Zwergtaucher |
| Greek | Νανοβουτηχτάρι |
| Gujarati | નાની ડૂબકી |
| Hebrew | טבלן גמדי |
| Hindi | छोटी पनडुब्बी |
| Hungarian | Kis vöcsök |
| Icelandic | Dverggoði |
| Indonesian | Titihan jelaga |
| Italian | Tuffetto |
| Japanese | カイツブリ |
| Kannada | ಗುಳುಮುಳುಕ |
| Korean | 논병아리 |
| Latvian | Mazais dūkuris |
| Lithuanian | Mažasis kragas |
| Malayalam | മുങ്ങാങ്കോഴി |
| Marathi | पाणडुबी |
| Mongolian | Хурган шунгуур |
| Norwegian | dvergdykker |
| Odia | ପାଣି ଡୁବୁକି |
| Persian | کشیم کوچک |
| Polish | perkozek |
| Portuguese (Angola) | Mergulhão-pequeno |
| Portuguese (Brazil) | mergulhão-pequeno-europeu |
| Portuguese (Portugal) | Mergulhão-pequeno |
| Punjabi (India) | ਡੁਬਕਣੀ |
| Romanian | Corcodel mic |
| Russian | Малая поганка |
| Serbian | Mali gnjurac |
| Slovak | potápka malá |
| Slovenian | Mali ponirek |
| Spanish | Zampullín Común |
| Spanish (Spain) | Zampullín común |
| Swedish | smådopping/indonesisk smådopping |
| Telugu | మునుగుడు కోడి |
| Thai | นกเป็ดผีเล็ก |
| Turkish | Küçük Batağan |
| Ukrainian | Пірникоза мала |
Tachybaptus ruficollis (Pallas, 1764)
Definitions
- TACHYBAPTUS
- ruficollis
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
25–29 cm; 130–236 g. Small and dumpy, the rump often appearing very fluffy. Nominate race breeding has blackish-brown cap down to just behind eye and to chin and upper throat, and back to hindneck, upperparts and upperwing-coverts; remiges brownish grey, with variable amount of white on secondaries and often also at base of inner remiges forming large panel on wing, the dark areas being paler on underwing except dark tertials, white underwing-coverts; chestnut on rear face (behind eye), lower throat, foreneck and broadly on sides of neck; dark brown from base of neck to flanks and breast, flanks fluffy and often show some chestnut and light buffy, whitish on abdomen and at base of tail, whiter on undertail-coverts; iris dark brown; bill black, with small whitish to yellowish tip, and obvious pale yellow to pale bluish basal patch ; legs dark greenish grey. Readily distinguished from other small grebes by chestnut lower throat and foreneck. Non-breeding adult is paler, upperparts dark grey-brown, with lower and rear face and most of neck buff to ochre-brown, rather than chestnut, chin and throat whitish, sides and flanks also paler, with much brownish to pale buff admixed; bill has orangey cutting edges and most of mandible, pale patch at base often greenish yellow. Leucistic individuals recorded, as are albinos (1). Sexes similar in plumage; as with most grebes, female marginally smaller than male. Juvenile similar to non-breeding adult, but usually retains some dark markings on face. Races differ mainly in size, in amount of white on secondaries, also in coloration: iraquensis is slightly smaller and darker than nominate, with less black on throat, white on inner secondaries, sooty suffusion below; <em>capensis</em> resembles previous, but iris yolk-yellow (variable, sometimes orange or dark brown), often with narrow pale line on anterior part of upper half of orbital ring, longer bill , also a leucistic/partial albino individual recorded in India (2); poggei has pale yellow eyes, rather long and slender bill (but smaller than in preceding race), restricted black on throat, only narrow pale tips to inner secondaries, silvery-white belly, and in non-breeding plumage exhibits less contrast between cap and face (especially as rear face and neck more strongly tinged brown); philippensis likewise has pale iris, perhaps also a pale yellowish eyering, and appears somewhat darker than last race; cotabato is mostly silvery whitish below; tricolor has long bill, plumage darker than others, mostly grey-black, with rufous ear-coverts and neck, and grey feather fringes to underparts; vulcanorum has more black on throat, but is variable, and only doubtfully distinct from last; collaris is similar to previous two, but lower neck black, and bill longer and heavier.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Closely related to T. novaehollandiae and †T. rufolavatus, both of which have been considered races of present species. Extensive hybridization with latter, and occasionally with T. pelzelnii; has interbred in SW England with vagrant Podilymbus podiceps, producing hybrid young (3). Internal taxonomy complicated and poorly understood; full revision required. Race tricolor sometimes considered a separate species (normally including collaris), on grounds of darker underparts and longer and more massive bill, as well as more pronounced sexual dimorphsim in bill length (4); however, although belly very much darker, bill appears only generally rather larger and no other differences apparent, so distinctiveness insufficient for separate species status. Race albescens has traditionally been included in capensis, but appears distinctive, especially with yellow vs brown iris (4). Races intergrade in some areas, e.g. albescens with poggei in Myanmar and W Thailand, cotabato perhaps with tricolor in zone from S Philippines to N Moluccas, and tricolor with collaris in New Guinea. Populations from Java and Lesser Sundas sometimes separated as race vulcanorum on basis of larger and more noticeable throat patch, but geographical reliability of this feature appears unclear (4). Race poggei includes proposed forms japonicus (Honshu) and kunikyonis (Ryukyu Is). Nine subspecies currently recognized.Subspecies
Little Grebe (Little) Tachybaptus ruficollis [ruficollis Group]




Distribution
Tachybaptus ruficollis ruficollis (Pallas, 1764)
Definitions
- TACHYBAPTUS
- ruficollis
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis iraquensis (Ticehurst, 1923)
Definitions
- TACHYBAPTUS
- ruficollis
- iraquensis
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis capensis (Salvadori, 1884)
Definitions
- TACHYBAPTUS
- ruficollis
- capense / capensis
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis albescens (Blanford, 1877)
Definitions
- TACHYBAPTUS
- ruficollis
- albescens
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis poggei (Reichenow, 1902)
Definitions
- TACHYBAPTUS
- ruficollis
- poggei
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis philippensis (Bonnaterre, 1790)
Definitions
- TACHYBAPTUS
- ruficollis
- philippense / philippensis
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis cotabato (Rand, 1948)
Definitions
- TACHYBAPTUS
- ruficollis
- cotabata / cotabato
The Key to Scientific Names
Legend Overview
Little Grebe (Tricolored) Tachybaptus ruficollis [tricolor Group]

Tachybaptus ruficollis vulcanorum (Rensch, 1929)
Definitions
- TACHYBAPTUS
- ruficollis
- vulcanorum
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis tricolor (Gray, 1861)
Definitions
- TACHYBAPTUS
- ruficollis
- tricolor / tricolora
The Key to Scientific Names
Legend Overview
Distribution
Tachybaptus ruficollis collaris (Mayr, 1945)
Definitions
- TACHYBAPTUS
- ruficollis
- collaris
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Wide range of wetlands, normally small and shallow, including small lakes, ponds, canals; also sheltered bays, vegetated shores of larger lakes and reservoirs. Outwith breeding season, occurs on more open waters; also, less frequently, on coast, in estuaries protected from large waves. In recent years, found also on newly created dams and sewage-farm ponds, especially in W Africa (5). From lowlands to mountains, in SW China to c. 4000 m; in Timor-Leste, race tricolor observed from sea-level to 2100 m (6).
Movement
Resident, dispersive or migratory, depending on winter temperatures on breeding grounds. Some in N of range (in E Europe especially) make post-breeding movement to tidal estuaries and sheltered bays, but generally far less common than other grebes in such habitats. Frequent records outwith main range, even in unsuitable habitat, indicate considerable capacity for aerial reconnaissance and colonization of new areas. Nominate race recorded in N Norway, Azores, Madeira and Canaries. In E of range, a rare migrant in Mongolia; autumn vagrant in L Baikal region (7).
Diet and Foraging
Mainly insects and larvae, especially mayflies (Ephemeroptera), stoneflies (Plecoptera), various water bugs (Heteroptera), beetles (Coleoptera), flies (Diptera), caddis flies (Trichoptera) and dragonfly larvae (Odonata); also molluscs, crustaceans and amphibians, mainly young, but also adults, e.g. small frogs , occasionally newts (Triturus); some small fish (e.g. of genera Cottus, Cyprinus, Gobio) taken. Largest prey are fish of up to 11 cm. Feeds mainly by diving for 10–25 seconds to depth of c. 1 m, rarely 2 m; also swims along with head and neck submerged, and picks prey directly off emergent vegetation or water surface. Sometimes associates with ducks and coots (Fulica) –which stir vegetation and flush small invertebrates – and on one occasion with scuba divers (8).
Sounds and Vocal Behavior
Loud, fast trill given by both sexes. In anxiety or alarm emits somewhat harsh “weeeib” notes and repeated short “pit” or “whit”.
Breeding
Season very variable, depending on growth of emergent vegetation and water level: Feb–Sept, with peak Apr–Jul, in W Palearctic and May–Jul (occasionally Oct–Feb) in Japan; recorded in all months in tropical Africa, and appears to breed throughout year in at least parts of Sundas (6); two broods, possibly three in warmer regions. Solitary. Nest a floating platform of aquatic plants anchored to submerged vegetation. Clutch 2–10 eggs, usually 3–5 (larger clutches perhaps product of two females); incubation period 20–25 days; chicks have dark down with pale stripes , paler below; fledging period 44–48 days, but young independent at 30–40 days. Success variable, in Europe 40% of hatchlings reach fledging stage; probably less than 1 young reared per nest in Africa. Oldest ringed individual 13 years of age.
Incubation
Conservation Status
Not globally threatened (Least Concern). Widespread and common in most of range. Global population estimated to lie in the range c. 610,000–3,500,000 individuals. In well-studied areas in Europe numbers fluctuate, mainly as a result of winter conditions: marked decline in Britain following cold winter of 1962–1963, and similarly in Netherlands after that of 1984–1985; increase in Scandinavia attributed to amelioration of climate during 20th century. Population in Britain and Ireland estimated at 9000–18,000 pairs in mid-1970s, with little evidence of subsequent change; in early 1990s, 6400–8000 in Germany, 7500–10,000 in Poland and 9000–10,000 in Hungary. Little information available on population sizes outside Europe: mid-winter counts in 1991 yielded 8450 in Kenya, 23,676 in India, 1359 in Pakistan, and 1035 in Japan; race iraquensis thought to number c. 6000 individuals; and capensis estimated at no more than 100,000 in Africa, with a similarly sized population in its Asian range. Negative effects of transformation of wetlands by destruction, pollution or recreational use are offset by construction of man-made ponds, reservoirs and dams, leading to expansion of species in many areas, e.g. E & S Africa. Has extended its range in recent decades: in C Arabia, irregular passage migrant and winter visitor in 1970s, increasingly more common during 1980s, bred for first time (at Al Hair dam) in 1990, and during 1990s a numerous breeder, although less common in 2003 than in 1999 (probably because R Riyadh had become choked with vegetation) (9); first bred on Ibiza (Balearic Is) in 2007 (10). In Madagascar has benefited from introduction of herbivorous fish, to the clear detriment of the two endemic species (T. pelzelnii, T. rufolavatus). Not always easy to survey, as breeding pairs can be overlooked in fringing reedbeds. Status in Borneo uncertain; has possibly bred (11).
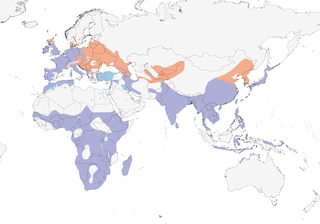
- Year-round
- Migration
- Breeding
- Non-Breeding

















